[Música] Hola cómo les va Bueno nada vuelvo a traer esto que veamos dejado cuando estuvimos hablando un poco de los procesos redox y es ver si ahora podemos encontrar cuál es la la pila que genera la energía para que las células puedan sobrevivir Así que vamos a hablar un poco sobre estos eventos de transferencia de electrones y la capacidad de síntesis de ATP vamos un poco por ahí básicamente en los procesos transductores de energía a nivel de la naturaleza encontramos dos grandes eventos que son la focalización oxidativa y las fotos fosforilación como eventos muy pero
muy relacionados con procesos muy similares Aunque en algunos este aspectos totalmente contrarios nosotros dado que vamos a estudiar a los seres humanos nos vamos a enfocar en la fosforilación oxidativa y vamos a tratar de poder entender cómo este proceso de transferencia de electrones que se da desde las coenzimas reducidas producidas a partir de las vías degradativas oxidativas que ya reconocemos al oxígeno como último aceptor de electrones termina siendo responsable de poder generar la suficiente cantidad de energía como para permitir la síntesis de ATP en el caso de la fosfotería Perdón fotoforilación que ocurre en los
organismos autótrofos y fácilmente básicamente aquellos organismos que tienen la capacidad de poder generar etp a través de la luz utilizan sistemas que son prácticamente similares nada más que en este caso y lo digo solamente como digamos como un como un comentario ellos utilizan agua y generan oxígeno como como producto sí un proceso que ocurre a nivel de los cloroplastos A diferencia de lo que ocurre en la configuración oxidativa a nivel de las mitocondrias y que de alguna manera tienen un origen bastante común posiblemente del punto de vista evolutivo vamos a hablar de la focalización oxidativa
como un proceso de síntesis de ATP impulsada por el proceso exergónico de transferencia de electrones desde las coenzimas reducidas al oxígeno sabemos que eso ocurre en las mitocondrias razón por la cual me parece que es bastante interesante hablar de algunos aspectos respecto de estas mitocondrias estos orgánulos de doble membrana con una membrana mitocondrial externa muy parecida a la membrana plasmática y una membrana metodolina interna este de una superficie mucho mayor por eso se generan las estructuras que reconocemos como crestas con características muy muy distintivas una membrana prácticamente impermeable A todo con una carga proteica este
realmente importante en un rato lo vamos a observar mitocondrias que como podemos observar en esta en este vídeo en donde las mito cuando eras coloreamos con un con una molécula que florece en verde hay en el enorme cantidad la digamos en una célula y que inclusive presenta dinámicas muy particulares si uno hace detalle en digamos en este vídeo en imagen Observa mitocondrias alargadas mitocondrias que se fusionan mitocondrias que se acortan hablando un poco de las mitocondrias en principio las mitocondrias los este las evidencias más fuertes definen su origen en un proceso endosimbionte en donde un
procarionte inferior un arqueobacteria posiblemente ha hecho a través de diferentes mecanismos sin necesidad de discutirlo la capacidad de generar una simbiosis con alguna otra arqueobacteria de tal manera que en ese proceso simbiótico Se generó este orgánulo que finalmente reconocemos como como mitocondria y que inclusive a partir de organismos fotosintéticos que también generaron simbiosis con alguna de estas células muy primitivas quizás los primeros esbozos de células eucariontes derivaron un li que dieron origen a lo que son las este las células vegetales que además de presentar mitocondrias presentan los cloroplastos hay enormes evidencias que demuestran esta teoría
en los simbiontes que van desde la particularidad de la doble membrana haciendo referencia posiblemente a la membrana propia de ese organismo original y la membrana del proceso de endocitosis la presencia de un ADN mitocondrial que es un ADN funcional es decir que presenta genes son 37 genes los que están codificados en el ADN mitocondrial genes que codifican para los arnés veintipico atrás ARN de transferencias un par de aviones ribosomales y para proteínas que conforman parte de lo que hoy vamos a estudiar como cadena respiratoria mitocondrial que ha sido muy útil en los procesos de tracking
y rastreo de evolución de humanos dado que la mitocondria básicamente es materna Y entonces se puede seguir a través de la secuencia del ADN mitocondrial los linajes en los humanos unas mitocondrias que son de una dinámica enorme no sólo un orgánulos estáticos sino que son orgánulos de una dinámica muy muy grande dado que pueden fusionarse pueden fusionarse en más mitocondrias dependiendo de las condiciones celulares esas mitocondrias puedan ingresar en apoptosis o te puede establecer la mitofagia y que inclusive esa mitocondrias van a estar ubicadas en lugares estratégicos en la célula dependiendo de la que la
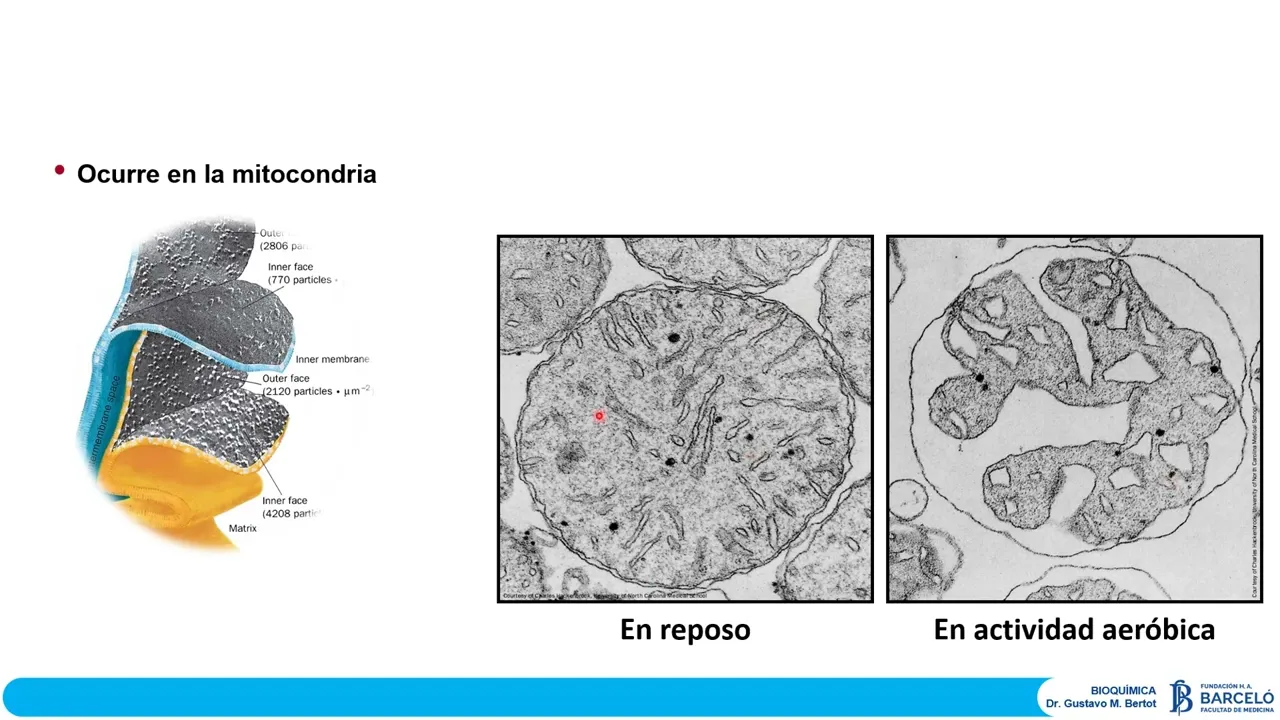
célula esté necesitando en momentos definidos y por supuesto mitocondrias que se pueden generar una de las características sobresalientes que tiene la mitocondrias y que inclusiven son fundamentales para poder entender un poco lo que estamos estudiando es analizar un poquito las características en la doble membrana que presenta esta membrana externa que observamos muy parecida a lo que es una membrana plasmática estándar y una membrana interna con características muy distintivas características que terminan definiendo las propiedades que van a ocurrir en esa membrana Y quizá los aspectos funcionales energéticos de la mitocondria más sobresalientes esto es una crío
fractura de la membrana externa y la membrana interna Esto está coloreado solamente para poder hacer diferenciación y lo que observamos acá como estructuras rugosas blanquecinas serían la presencia de proteínas presentes en esas membranas y como ustedes pueden ver acá en esta imagen la membrana mitocondrial interna en su capa más del lado de la matriz tiene una carga de partículas por unidad de área que es prácticamente el doble de lo que se encuentra en la membrana interna pero en su fase digamos cercana al espacio intermembrana y muchísimo muchísimo mayor de lo que es una distribución de
proteínas que encontramos en la membrana mitocondria externa de tal manera que entonces esta membrana mitocondría interna es una membrana Ultra proteica más del 80% de esta membrana es proteína y la define como una membrana quizás la membrana más impermeable a todo básicamente a través de esta membrana va a pasar agua va a pasar cualquier tipo de gas pero va a estar restringido salvo por la presencia de transportadores específicos o inespecíficos pero transportadores o carnes al fin la posibilidad de intercambio que pueda haber entre la mitocondria y su entorno otra de las características sobresalientes respecto de
de situaciones estructurales en esta membrana mitocondrial interna es la presencia de este fosfolípido un fosfolípido que no encontramos en ninguna otra membrana que no sea esta pero que sí encontramos en las membranas de los procariontes un dato más que avala la teoría en los simbiontes que es la cardiolipina la cardiolipena es un fosfolípido que presenta una estructura bastante particular dado que si uno reconoce a los fosfolípidos como estructuras anfipáticas con dos con una cabeza polar y dos colas no polares la cardiolipina presenta cuatro colas no polares y Define papeles fundamentales en esa mitocondria que van
desde regulación de la apoptosis los procesos de difusión y fisión mitocondrial respuestas de señalización a moléculas Y fluidez de esa membrana fíjense lo que ocurre cuando a una membrana estándar le voy agregando cardiolipina como un fosfolípido distintivo fíjense como por la estructura que presenta esta cardiolipina obliga esa membrana a generar estructuras este diferentes de tipo circulares y con ciertos plegamientos Y quizá es hacia parte de la razón explica por qué la membrana mitocondrial interna es una membrana que presenta esos plegamientos que son característicos que nosotros denominamos creencias algo que es muy interesante es lo que
le presenta nuestra en esta diapos esta es la mitocondria de músculo esquelético en una condición de reposo y su correlato en lo que sería una misma mitocondria del mismo músculo pero en actividad hay claras diferencias estructurales entre esa mitocondria y un músculo en reposo versus esa mitocondria en un músculo en actividad y si uno puede reconocer esa estructuras se da cuenta que lo que está ocurriendo es que o hay una contracción de la matriz mitocondrial o hay una expansión del espacio intermembrana entonces acá se plantea una serie de preguntas y si es una contracción de
la matriz matrimonio mitocondrial Cuáles son los fenómenos asociados a la actividad que llevaron a esta contracción Y si la postura es que hay una expansión del volumen del espacio intermembrana La pregunta es qué es lo que hay dentro de ese espacio intermembrana que obligas a esta expansión y en consecuencia esta supuesta contracción de la matriz mitocondrial Se los voy dejando como inquietud así a ver a medida que vamos desarrollando podemos encontrar digamos respuestas a estas dos posibles situaciones dijimos que la fosforilación oxidativa se refiere a la síntesis química de ATP conducida por el proceso exergónico
de transferencia de electrones desde las coenzimas reducidas al oxígeno acabamos de comentar su ocurrencia en la mitocondria con algunos aspectos fundamentales respecto de la mitocondria ahora vamos a hablar de la fosforilación oxidativa como un proceso que comienza con el ingreso de electrones a lo que definimos como cadena respiratoria o cadena del transporte de electrones en donde los electrones pasan a través de una serie transportadores que encontramos incluidos en la membrana interna mitocondrial vamos a hablar un poco de estos transportadores electrónicos o vamos a volver a referirnos algunos de estos transportadores electrónicos estos son los cinco
grandes transportadores electrónicos que vamos a encontrar asociados a esta cadena de transporte de electrones observamos algunas especies que son ya conocidas por nosotros otras quizás no tanto encontramos [Música] y al fmn a la coenzima q a una familia demo proteínas que son los citocromos y una familia de proteínas muy muy ansentes muy muy viejas en la escala biológica que son proteínas que contienen centros con azufre y con hierro vamos a analizar un poquito cada una ya hemos comentado sabemos que Son estructuras químicas que tienen la capacidad de poder participar de los procesos de transferencia de
electrones y que inclusive cada una lo hace con ciertas particularidad Mientras que el Nat y el Nat p tiene la capacidad de poder transferir electrones de a dos en bloque el fácil fmn lo pueden hacer de a uno de tal manera de generar estructuras intermedias semiquinonas llamadas radicales respecto de la coenzima q algo que es interesante remarcar es que esta coenzima q tiene toda esta cola isopreno esta cola de 10 isoprenos le da carácter lipídico a esta a esta molécula por encima cubo o ubiquinona también es nombre conocido es interesante porque Define su ubicación como
parte de la membrana y que tiene la particularidad de poder participar de procesos de transferencia de electrones de manera tal de poder transferir dos electrones en forma completa desde su estado totalmente oxidado has estado totalmente reducido pero de a 1 por mes de manera similar a lo que ocurría con el fmn y el fado Y encontrándose nuevamente un estadio intermedio como radical es decir con un electrón desapareado que va a tener interesantes consecuencias para charlar más más adelante tenemos a los hipocromos que son estas hemoproteínas que dependiendo de el tipo de emo y como se
asocia con la parte proteica definirá toda una familia de citocromos que reconocemos con letras ABC y subtipos entre el caso y en donde el hierro es el el punto central de los procesos de transferencia de electrones ya hemos comentado esta característica que presenta el hierro de poder pasar de hierro oxidado de hierro reducido Y en este sentido permitir la transferencia de un electrón y finalmente las proteínas con hierro y azufre o sulfo hierro proteínas una familia bastante amplia de proteínas que tiene como características central tener grupos protéticos conformados por núcleos de azufre Unidos a hierro
con distintas estructuras y que de la misma manera que podemos reconocer en los citocromos al hierro como el elemento que participa de estos procesos redox indudablemente en estos en estas proteínas con hierro de azufre el hierro cumple la misma función es la manera que Entonces estamos encontrando especies químicas que tiene la capacidad de poder transferir electrones de a dos en bloque de a uno o de a dos pero pudieron darse secuencialmente de a uno volviendo algunos conceptos que hemos charlado Nosotros sabemos que podemos definir el flujo en un proceso de transferencia de electrones reconociendo los
potenciales redos que presentan las especies de forma tal que sabiendo que los electrones fluyen desde los potenciales más negativos hacia los potenciales más positivos el reconocimiento de cada uno de estos transportadores electrónicos permitió establecer una secuencia de flujo entre ellos desde las coenzimas reducidas como los dadores de electrones hacia el oxígeno como el gran aceptor de electrones pasan Entonces por toda por toda esta serie de transportadores electrónicos en una secuencia definida y determinada por sus potenciales redox que establecen el orden en el que uno que va a poder encontrarlos a nivel de la membrana mitocondrial
interna por otro lado sabemos que estos procesos de transferencia de electrones están claramente asociados a procesos de cambios en la energía libre del sistema razón por la cual una nueva pregunta para hacernos es si a nivel de la membrana mitocondria interna estos pasajes de electrones que se están dando a través de estas especies no podrán generar energía suficiente para poder recuperar bajo la forma de ATP hicimos entonces que la fosforilación oxidativa se refiere a la síntesis química ATP conducida por el proceso excergónico de transferencia de electrones desde las coenzimas reducidas al oxígeno sabemos que es
el mitocondrial sabemos que está implicada un proceso de cadena de transporte de electrones definimos que esos electrones pasan a través de una serie de transportadores que estimamos que van a estar incluidos en la membrana de mitocondrial interna y que en realidad esos transportados electrónicos funcionan dentro de complejos proteicos que están ordenados en serie para poder cumplir con esta situación de transferencia de acuerdo a sus potenciales redox vamos a hablar entonces un poco de los famosos complejos que conforman la cadena respiratoria básicamente hablamos de cuatro complejos así fue como fueron definidos independientemente que un complejo pueda
contener más de una proteína complejo 1 complejo 2 complejo 3 y complejo 4 en donde como podemos observar en esta imagen encontramos incluido a estos transportadores universales de electrones como son las proteínas con hierro y azufre o enzimas como el flavismo mononucleótido citocromos encontramos en estos complejos y vale la pena remarcar en este caso la presencia de la coenzima q no asociado a ninguno de los complejos dado que como habíamos comentado su característica lipídica le permite poder fluir libremente en esta membrana mitocondrial interna otro punto para remarcar respecto de complejos o proteínas que conforman parte
de esta cadena el transporte de electrones es la presencia de una proteína que contiene el citocromo c que como este esquema lo indica fíjense que parece estar asociada a la membrana mitocondrial este interna en su lado del espacio intermembrana pero de una manera bastante libre de tal manera que uno puede predecir que tenemos el complejo 1 el complejo 2 el complejo 3 y el complejo 4 embebidos en la membrana mientras que la coenzima q y el citocromo c parece intercambiadores móviles que pueden potencialmente conectar estos complejos entre sí de tal manera que entonces el complejo
1 el complejo 2 el complejo 3 y el complejo 4 son las estructuras proteicas que contienen a los transportadores electrónicos de tal manera de permitir el flujo de los mismos desde las coenzimas reducidas pasando secuencial por estos complejos hasta el oxígeno para poder conformar este evento que conocemos como cadena respiratoria vamos a hablar un poco sobre sobre esto es interesante que cuando estudiaron la presencia de estos de estas proteínas como parte de la membrana mitocondrial interna además de estos cuatro complejos que participaban en la transferencia de electrones encontraron un quinto complejo del cual hablaremos más
tarde era o mejor dicho que es el complejo Que está más asociado a la síntesis de ATP una nueva pregunta para hacernos es si Existe algún tipo de conexión entre los cuatro complejos que conforman la cadena respiratoria es decir la cadena de transporte de electrones y el complejo 5 llamado tp-sintasa que es el complejo encargado de la síntesis de ATP en un esquema un poco más este simplificado que es el que va a aparecer posteriormente en las diapositivas siempre siempre tener referencia de la ubicación de la membrana mitocondrial interna el espacio intermembrana la matriz mitocondrial
y en tal caso por acá arriba la membrana metocondrial externa para poder referirnos al complejo 1 al complejo 2 al complejo 3 al complejo 4 y como mencionábamos hace un ratito la coenzima q como un lípido móvil y el citocromo c también como una proteína que al estar débilmente unida a la membrana también participa de procesos móviles el complejo 1 o También conocido como nada H deshidrogenasa Es por donde ingresan los nada H provenientes de las vías oxidativas tanto mitocondriales como aquellos nadachi que puedan proceder de transferencia desde el citosol Y en donde vamos a
encontrar toda una serie de proteínas con hierro y azufre flavin mononucleótidos de tal manera que los dos electrones que el nadache esta sería nuestro proceso de oxidación son transferidos en forma secuencial hacia la coenzima q como la coenzima encargada de poder recepcionarlos vemos en un vídeo solamente a nivel ilustrativo de lo que está sucediendo hayamos representado nuestro ya conocido ciclo de krebs y vamos a ver cómo tomamos una de estas reacciones características del ciclo de krebs que es la transformación de malato en oxalato por la mandato de hidrógenasa que produce nada H como coenzima redox
acá estamos en presencia de la mandato deshidrogenasa que genera el nadache como parte del producto y que como Nosotros sabemos va a ir a reoxidarse al complejo 1 de la cadena respiratoria y a los que estamos observando en esta imagen fíjense que por la membrana mitocondrial están apareciendo unas moléculas móviles después como vamos a revisarlas mejor que son las coenzimas y fíjense como esta animación nos va mostrando la disposición de por un lado el flamismo mononucleótido y los centros de proteínas con hierro y azufre de tal manera que estos dos electrones que el nadache está
transfiriendo al flavismo mononucleótido van a ser transferidos en forma secuencial a lo largo de toda este brazo del complejo 1 a través de los complejos con proteínas con hierro y azufre de tal manera de poder ir cargando de electrones a la coenzima Q que Recuerden que tienen la capacidad de poder ir tomándolo de a uno hasta llenarse de dos y dada su capacidad de ser un lípido este tiene la posibilidad de poder movilizarse a través la membrana si ustedes observan existen cuatro puntos en este caso círculos rojos que corresponde nada protones y fíjense que en
este proceso de transferencia de electrones hay un posicionamiento particular de protones sobre ciertas regiones del complejo de tal manera que en un momento determinado esos protones ubicados Sí en la matriz mitocondrial terminan siendo transferidos hacia el espacio intermembrano es decir que uno podría resumir la característica de este complejo 1 como una proteína que permite por un lado la transferencia de electrones desde el nadache hacia la coenzima q de tal manera que la coenzima q transferiría a sus electrones posteriormente es alguna especie y que al mismo tiempo en este proceso hay una ubicación particular de protones
que terminan siendo transferidos desde la mitocondria desde la matriz mitocondrial perdieron al espacio intermembrano el complejo 2 es succionato deshidrogenasa y cuando anís el suxinato de hidrogenasa piensa estamos hablando de krebs y La respuesta es sí estamos hablando de crepes la solucinato es hidrogenasa Es una de las enzimas del ciclo de krebs y tiene una ubicación particular que es en vez de ubicarse en la matriz mitocondrial la encontramos unida a la membrana mitocondrial interna la razón que pueda explicar esta ubicación tan definida es que contiene fat como grupo prostético y dado que el fa de
es un grupo prostítico y no tiene capacidad de estar libre como lo hizo el nadache el proceso de reoxidación del fat cuando transforma el suxinato en fumarato requiere una transferencia directa a una especie que se encuentra en la membrana mitocondrial interna razón por la cual su ubicación es estratégica para poder Entonces cerrarles los electrones a la coenzima q dentro de esta succionato es hidrogenasa también encontramos sin formando parte del complejo algunos centros de proteínas con hierro de azufre de tal manera que estas electrones transferidos del succionato al fumarato en esta secuencia que hemos visto del
ciclo de krebs terminan siendo transferidos a la ubiquinona Y entonces la ubiquinona o la coenzima q Parece ser una especie particular que permite la recepción de electrones no solamente el complejo 1 sino también del complejo 2 e inclusive de cualquier otro conjunto de proteínas y al tener faz como grupo prostético deben ubicarse estratégicamente sin la cerca encima q para poder transferirle esos electrones el complejo 3 es También conocido como citocromos ser reductasa y acá es donde empezamos a encontrar a los citocromos como parte de especies de transferencia de electrones es la coenzimacula responsable de transferir
los electrones que provienen o del complejo 1 o del complejo 2 al complejo 3 dentro del complejo 3 encontramos además de centros con hierro y azufre una serie de citocromos sí que sin ser muy puntuales vamos a encontrar el citocromo según el citocromo B subtipos de citocromode lo interesante es poder referenciar que acá estamos en un proceso en donde hay un transportador electrónico que es la coenzima Q que tiene la capacidad total de transferir los electrones pero que las especies internas dentro del citocromo C3 reductaso del complejo 3 tiene la capacidad de poder tomar solamente
de a uno es decir que la tiene que poder vaciar este transporte de electrones y es así que ustedes Cuando lean en los textos van a ver que a este evento que ocurre en el complejo 3 se lo conoce como el ciclo de la coenzima q o ciclo q dado que la coenzima q va a entregar de a un electrón en forma independiente a distintos citocromos de tal manera que en este juego de entrega de electrones se terminan conformando la transferencia de 2 a un electrón desde la coenzima q al citocromo C fíjese que nuevamente
este proceso de transferencia de electrones desde la coenzima q al citocromo c a través del complejo 3 nos muestra sí asociación con el pasaje de protones desde la matriz mitocondrial al espacio intermembrano final mente el complejo 4 también llamado citocromo se oxidasa ositocromo a A3 dado que en su estructura encontramos un citocromo a y un citocromo A3 asociados ambos a centros con cobre uno de los aspectos interesantes de los procesos de transferencia de electrones que ocurren a nivel del complejo 4 provenientes del citocromo C es la presencia del oxígeno que tiene la capacidad de poder
tomar los cuatro electrones para reducirse completamente agua pero poder hacerlo de a uno de ahí la importancia que tienen estos dos centros de digamos los dos centros hemoa y hemos A3 asociados al cobre para poder permitir este juego electrónico de transferencia en forma secuencial de tal manera de al mismo tiempo que se están transfiriendo estos electrones nuevamente aparece un transporte de protones Asociados la cadena de transporte de electrones permite entonces la reacción de las coenzimas reducidas que fueron productos de los procesos metabólicos de oxidación de las biomoléculas por otro lado la cadena de transporte de
electrones nos muestra que es un proceso exergónico que libera energía suficiente para la síntesis de ATP algo que hemos ya revisado más de una vez la transferencia de dos electrones desde el nadache al oxígeno libera 52 kilocalorías por mol es decir en este proceso redox que ocurre a nivel de la cadena respiratoria se está liberando suficiente energía como para poder predecir la síntesis de ATP La pregunta es De qué manera esta pila logra conectarse con los sistemas generadores de energía ahí aparece la ATP sintasa el complejo 5 que es lo que vamos a analizar a
continuación el ATP es conocido desde 1930 la relación entre ATP y respiración se conoce desde los años 40 en 1961 Efraín racker aísla parte de lo que conocemos actualmente como la ATP sintasa aísla el componente f1 de la tp-sintasa a través de un procedimiento relativamente este habitual para lo que eran los estudios en ese momento de orgánulos que fue aislar mitocondrias y eres aislamiento mitocondrial permitir que la membrana mitocondrial interna quedará expuesta y que entonces a partir de esta exposición estas estructuras que ellos observaban a nivel de la microscopía electrónica como protuberancias que se observaban
en la membrana de mitocondría externa quedarán expuestas y a través del uso de detergentes poder separarlas de esa membrana así fue como racker logró aislar estas componentes y que comenzó a relacionar como estructuras asociadas a la generación de energía contemporáneo digamos a Raquel en los años 60 como vamos a ver ahora digamos a continuación ya empezaban a postularse cuáles eran los mecanismos a través del cual esa cadena de transporte de electrones se conectaba con lo que ya se empezaba a conocer que era la actividad de esta proteína con la capacidad de síntesis de ATP en
1966 Raquel hace un experimento que es fundamental para la comprensión del funcionamiento de esta de esta proteína ya con el conocimiento de lo que Peter mitchell había postulado también en los años 60 el ya tenía aislada la ATP sintasa en forma completa Y a partir de las teorías de Míchel que ahora lo comentamos a continuación postulaban de una fuerza dominada por los protones él logra construir un sistema artificial para demostrar Cómo es que esta proteína funcionaba en la isla una proteína este bacteriana que actúa como un canal para protones y ese canal para protones se
estimula por la luz de tal manera que él construyó un compartimento liposomal en donde ancló a esta bomba de protones bacterianas estimulada por la luz y ancló también a su proteína en estudio que era la de síntesis de ATP Porque en ese momento el postulado era que la síntesis de ATP tenía que ver con gradientes de protones y básicamente lo que hizo fue en este sistema artificial conformado por esta bacteria rodoxina estimulada por por la luz que funcionaba como un canal para protones el demostró que cuando estimulaba con la luz y es de esa manera
se abría este canal este ingreso de protones al espacio interno de este Roma era necesario para permitir la síntesis de ATP esto lo observó en un simple gráfico Y es que a medida que iluminaba a su sistema su sistema generaba ATP la solidación oxidativa dijimos que se refiere a la síntesis química de ATP conducida por el proceso extergónico de transferencia de electrones desde las coenzimas reducidas al oxígeno que ocurre en la mitocondria que los electrones pasen a través de una serie transportadores incluidos en la membrana interna que esos transportadores electrónicos funcionan dentro de complejos proteicos
ordenados en serie complejo uno complejo 2 complejo 3 complejo 4 y a su vez se encuentran conectados por la coenzima q y el citocromo c como transportadores móviles que esta cadena de transporte de electrones es un proceso exergónico que libera energía suficiente para la síntesis de ATP y que existe una translocación de protones desde el espacio desde la matriz mitocondrial al espacio intermembrano que de alguna manera fuimos definiendo a medida que fuimos analizando estos procesos de transferencia de electrones cada vez que nosotros observamos el pasaje de electrones hacíamos mención en el complejo 1 del complejo
3 y en el complejo 4 de que estos pasajes de electrones implicaban también el pasaje de protones desde la matriz mitocondrial al espacio intermembrana hicimos referencia a cómo en el complejo 1 cuando este brazo se cargaba de electrones eso empujaba a protones ubicados en sitios específicos a que sean transferidos de la matriz al espacio intermembrana hemos comentado Cómo la coenzima q a través de su capacidad de poder transferir y a un electrón permita que existieran transferencias de protones desde la matriz mitocondrial al espacio intermembrana y hemos comentado cómo esos procesos de transferencias secuenciales de electrones
desde el citocromo al citocromo A3 utilizando los complejos asociados al cobre también permitía la apertura de canales de transferencia de protones de tal manera que lo que nosotros estamos observando es que a medida que ocurre la transferencia de electrones se produce el bombeo específico de protones a nivel de complejo 1 a nivel del complejo 3 y a nivel de complejo 4 y definimos un veo porque habíamos referido al principio que la membrana mitocondrial interna es una membrana impermeable prácticamente a toda molécula salvo agua y gases en 1961 Peter mitchell postula La teoría quimiosmótica mitocondrial Recuerden
que en 1961 ya racker había logrado aislar la f1 la fracción f1 de lo que ahora conocemos como la ATP sintaxia Estos son este es uno de sus trabajos originales Este es el digamos el título de otro de sus trabajos originales en donde él comienza a referir y a postular De qué manera es que se sintetiza tpu la cinta CD a nivel mitocondrial tuvo a los bioquímicos durante mucho tiempo en vilo en función de que todos apostaban a que la ATP se sintetiza de una manera química clásica es decir a través de una reacción en
donde estequiométricamente las cosas resultaban de desde un punto de vista químico muy sencillo tanta cantidad de B más tanta cantidad de c la tanta cantidad de X ellos observaban que cuando analizaban los rendimientos energéticos no seguían las leyes de la química y entonces algún grupo entre los que se encontraba Michelle entre los que se encontraban trackers entre lo que se encontraban boyle este y otros investigadores empezaron a observar algunos otros eventos de la naturaleza en donde la energía funcionaba a través de la generación de gradientes Y esto es lo que Michelle postuló como parte de
lo que se conoce como la teotimismática que termina siendo en realidad la forma en que la que la naturaleza diseña la estrategia de síntesis de ATP a nivel mitocondrial miche lo que postula es que se genera un gradiente de protones a nivel del espacio intermembrana y que este gradiente de protones genera otra vez del pasaje de electrones por los complejos 1 2 3 4 genera una fuerza protomotriz es decir una fuerza dominada por un cambio de pH que hay entre la matriz y el espacio intermembrana por digamos la transferencia de protones y es esa fuerza
protomotriz la que puede ser Entonces recuperada como energía bajo la forma de ATP los postulados De mitchell nada esto es solamente a modo informativo definían que el transporte de electrones a través de la cadena respiratorias se encuentra social transporte de protones desde la matriz al espacio intermembrana que la membrana mitocondria interna es impermeable a los protones de tal manera que Entonces digamos se genera un gradiente real que se conserva la energía de oxidación de esos procesos metabólicos en forma de un potencial electroquímico en la generación de un gradiente electroquímica de protones y que esa cadena
de transporte de electrones se encuentra acoplada a la síntesis de ATP el flujo de protones a favor de su gradiente electroquímico proporciona entonces la energía libre necesaria para la síntesis de ATP es decir que estos protones que son bombeados desde la matriz mitocondrial a nivel del complejo 1 a nivel del complejo 3 y a nivel de complejo 4 vuelven únicamente a través de la ATP sintasa y es esta vuelta a favor de ese gradiente lo que permite la utilización de la energía para la síntesis de ATP este potencial químico esta Delta de pH que se
establece entre la matriz y el espacio intermembrana este potencial eléctrico Dado que los protones tienen carga negativa son el fundamento que Define a la famosa fuerza protón motriz que es la que conduce la síntesis de ATP volviendo a esta día pues donde nosotros observamos esta característica distintiva en dinámica de una mitocondria y un músculo en reposo versus la misma mitocondria del músculo en actividad Ahora sí podemos reconocer la razón por la cual cuando este músculo entra en actividad encontramos Este cambio tan marcado a nivel de la matriz y del espacio intermembrano podemos concluir entonces que
no es que hay una contracción de la matriz mitocondrial sino que en este proceso de actividad muscular lo que estamos observando es como esta transferencia de electrones genera este gradiente de protones que va cargando y va llenando al espacio intermembrana de protones porque son esos protones lo que entonces la mitocondria utilizará para poder sintetizar ATP es decir que lo que está de alguna manera llenando todo este espacio que observamos acá en blanco son protones en 1997 polvoyer y John Walker reciben el premio Nobel en función de sus trabajos sobre la ATP sin tasa la te
presentaste es este complejo proteico conformado básicamente por dos componentes este componente que protruye que es lo que Raquel purificó en los años 60 que aparecen las micro fotografías electrónicas como una prótesis protuberancia tipo bolita que está conectada a través de un tallo con una estructura llamada f0 o mejor dicho ffo o de oligomicina dado que esta subunidad es sensible a el antibiótico oligomicina y Qué es finalmente la estructura proteica encargada de la síntesis de ATP de tal manera que los protones acumulados en el espacio intermembrana son transferidos a través de las unidades fo y en
esa transferencia se producen cambios conformacionales que permitan Entonces a las unidades f1 actuar como encima de síntesis de ATP el gradiente de protones la energía contenidas de gradiente de protones es la energía que utiliza la atpasa para poder sintetizar ATP ellos demostraron parte de su premio Nobel tiene que ver con este experimento un experimento es interesantísimo en donde ellos demuestran que esta encima trabaja como si fuera un motor molecular la imagen que estamos observando es una imagen original de sus experimentos en donde ellos anclan a la subunidad f1 y el tallo a un filamento de
actina hacen hacen funcionar a través de un sistema generador de protones y eso se observan como digamos la molécula gira a través de su eje y la única explicación posible a este movimiento circular es que indudablemente esa molécula esté funcionando como si fuera un rotor de tal manera que ellos observan que al mismo tiempo estas unidades f1 tiene la capacidad de poder trabajar en tres fases en forma continua dado que tiene tres sitios un sitio para poder digamos un momento Mejor dicho un momento en donde une adp más fósforo otro momento en donde se está
formando el ATP y un tercer momento en donde la tp es liberado sí lo podemos observar en este esquema representativo como las unidades f0 funciona como un rotor que gira como consecuencia del pasaje de protones del espacio intermembrana a la matriz mitocondrial y como ese giro de ese rotor 9 sin el tallo que altera la estructura de las subunidad f1 de tal manera de permitir al mismo tiempo esos tres procesos que estábamos comentando para la síntesis de ATP Entonces cuando nosotros analizamos un poco en forma global este proceso que ocurre a nivel de la mitocondria
observamos como este proceso exergónico que ocurre como consecuencia de la transferencia de electrones a través de los complejos presentes en los que denominamos cadena respiratoria genera la energía suficiente para poder bombear protones y quedada la impermeabilidad presente en la membrana mitocondrial interna se genera un gradiente electroquímico de protones esa fuerza protomotriz que es la que la ATP sintasa utiliza para poder conducir un proceso endergónico como es la síntesis de ATP analizando algo mucho más cotidiano podemos encontrar como nosotros humanos hemos aprovechado el almacenamiento de agua a través de un dique con tener un montón de
energía potencial que puede ser utilizada cuando el agua es transferida a través de conductos determinados que permiten el movimiento de una turbina y que esa turbina a través su rotor hace Girar un electroimán y es ese electroimán el responsable de poder transformar toda esa energía cinética que se obtiene por el movimiento de la turbina en energía eléctrica interesante esta comparación entre la estrategia de obtención de energía que tiene una mitocondria a través de la generación de un gradiente y la obtención de energía en una central hidroeléctrica a través de la acumulación de energía bajo la
forma de agua y el movimiento de motores en un caso la turbina que mueve el electroimán para transformar la energía cinética del agua en energía eléctrica y el motor molecular del ATP sintasa que funciona en este caso a protones y en donde ese flujo de protones mueve ese rotor que permite En consecuencia transformar esa energía contenida en el gradiente en ATP la moneda que la célula utiliza para poder conducir todos esos procesos endergónicos con esto entonces finalizamos un poco esta revisión sobre cadena respiratoria y fosforilación oxidativa y dejamos entonces para análisis seguramente en clase dado
que es mucho más interesante hacerlo desde el punto de vista sobre desacoplantes e inhibidores de la cadena respiratoria y de la fosforilación oxidativa tratando de hacer foco en Cuáles son los efectos de los acoplantes Cuáles son los efectos de los inhibidores ejemplos de desacoplantes fisiológicos y algunos ejemplos de inhibidores digamos que no son fisiológicos Pero los que podemos estar expuestos los dejo Cuídense hasta la próxima















![Upbeat Lofi - Deep Focus & Energy for Work [R&B, Neo Soul, Lofi Hiphop]](https://img.youtube.com/vi/THh4fT0O7IY/maxresdefault.jpg)



![Instant Energy Boost ☕️ [upbeat lofi / productivity mix]](https://img.youtube.com/vi/LhCXGVW8-CU/maxresdefault.jpg)