Lezione 22. I mitocondri sono organelli addetti alla respirazione cellulare. Essi presenti praticamente in tutte le cellule eucariote.
Ogni cellula mediamente ne contiene 2000. Questo numero però non è fisso, ma può variare da cellula a cellula in base alle differenze di richiesta energetica. Una cellula muscolare avrà molti mitocondri, molti di più rispetto a quelli di una cellula epiteliale.
Inoltre, il numero può variare anche all'interno di una stessa cellula, a differenza dello stato funzionale. Una cellula può alternare fasi di alta attività e lunghi periodi di bassa attività in cui non necessita molta energia e per questo il numero di mitocondri si riduce. Il numero di mitocondri, inoltre, non è uniforme in tutto il citoplasma e varia con la disposizione.
Infatti nel negli spermatozoi i mitocondri si accumulano principalmente attorno al flagello, che è la struttura che richiede più energia. Il resto del citoplasma invece ne è praticamente vuoto. Il mitocondrio è composto da una struttura a due membrane.
Quella esterna separa il citoplasma dallo spazio intermembrana, quello spazio che si trova tra le due membrane. Questa è del tutto simile a una membrana citoplasmatica. Questa contiene infatti colesterolo e un rapporto lipid proteine del 50%.
Questa membrana contiene inoltre numerose acquaporine che la rendono fortemente permeabile. Poi vi è lo spazio intermembrana, quello spazio che separa la membrana esterna e la membrana interna. Questo è essenziale per creare il gradiente protonico durante la fosforillazione ossidativa, ma vedremo questo più avanti.
La membrana interna invece non ha la semplice funzione di delimitare la struttura, ma è ricca di proteine, proteine che svolgono la fosforilazione ossidativa. Il rapporto proteine lipidi è infatti del di 3 a 1, quindi ci sono numerosissime proteine concentrate. Un'altra caratteristica di questa membrana, che la differisce da quella esterna e dalla membrana acitoplasmatica della cellula è l'assenza di colesterolo e di acqua porine.
Questa membrana, inoltre, non è lineare, ma è ripiegata. Questo serve ad aumentare la sua superficie e permettere maggiori scambi durante la fosforillazione ossidativa. All'interno di questa membrana vi è la matrice mitocondriale.
In questa matrice ci sono numerosi enzimi proteici e ribozzimi, tra cui i ribosomi. Inoltre, all'interno di questa matrice vi è del DNA, un DNA mitocondriale. Il DNA del mitocondrio però è diverso da quello della cellula di cui fa parte.
Questo DNA, innanzitutto è un DNA a doppio filamento ad anello. Questo, inoltre, non si organizza in cromosomi e quindi non è avvolto attorno a degli stoni. È un DNA inoltre molto corto e presenta due geni per gli RNA ribosomiali, 22 geni per gli RNA transfer e 13 geni per produrre gli enzimi fondamentali per la fosforilazione ossidativa.
La presenza di questo DNA è importante perché permette al mitocondrio di replicarsi autonomamente, si comporta come se fosse una cellula all'interno della cellula. Inoltre, questo DNA si trasmette per via matrilineare perché al momento della fecondazione i mitocondri presenti all'interno dello spermatozzo vengono persi. La presenza di un DNA circolare e la struttura a doppia membrana viene spiegata dalla teoria dell'endosimbiosi.
Secondo questa teoria il mitocondrio era una cellula procariote che è stata fagocitata da una cellula eucariote. Questa cellula eucariote però invece di degradare il fagosoma, ha sviluppato con esso un rapporto di simbiosi. La cellula eucariote fornisce precursori per i processi e la cellula procariote, in risposta produce energia.
Questo è vantaggioso per entrambe le cellule. La procariote riceve numerosi precursori e per produce tanta energia. Questa energia, oltre a permettere la sua sopravvivenza, permette la sopravvivenza anche della cellula eucariote.
Ovviamente, essendo una cellula autonoma all'interno di un'altra cellula, il mitocondrio può duplicarsi in modo autonomo utilizzando il suo DNA ed i suoi enzimi. Come abbiamo accennato, la funzione principale del mitocondrio è quella di produzione di energia, ovvero il metabolismo cellulare. Il metabolismo attuato dai mitocondri trasforma molecole semplici come zuccheri, amminoacidi e acidi grassi in energia.
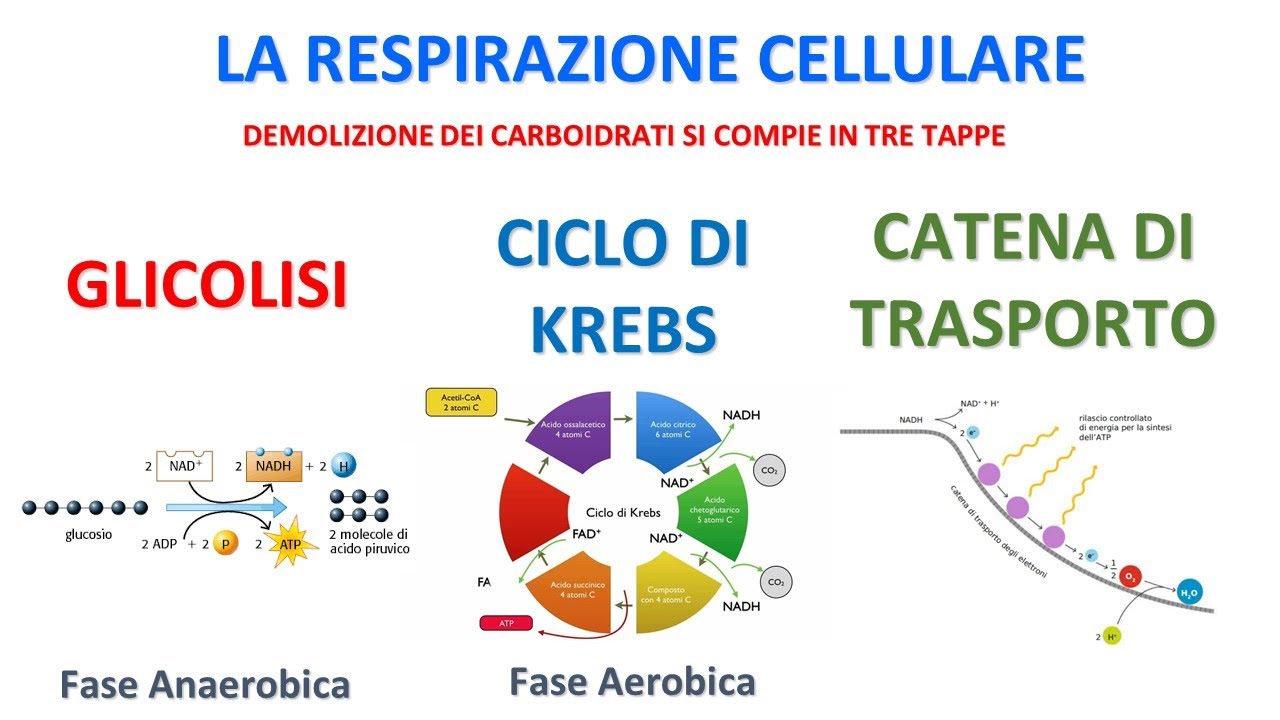
Questa energia verrà prodotta sotto forma di ATP. Per comprendere il funzionamento di mitocondri osserveremo in questa lezione il metabolismo del glucosio, un monosaccaride. Il glucosio all'interno del citoplasma della cellula va incontro alla glicolisi, ovvero alla trasformazione in due molecole più semplici.
A questo punto l'attività del mitocondrio si avvierà solo se c'è dell'ossigeno a disposizione. Per questo motivo il metabolismo dei mitocondri detta anche respirazione cellulare e questo avverrà in diverse fasi. Se non c'è a disposizione l'ossigeno, i prodotti del glucosio andranno incontro alla fermentazione.
Osserviamo ora queste singole tappe nello specifico. La prima fase della glicolisi è la fase di investimento. In questa fase la cellula spende due molecole di ATP per scomporre il glucosio in due molecole più semplici.
La repulsione dei gruppi fosfato. raggiunti al glucosio, infatti, lo spezzano in due molecole più semplici a tre atomi di carbonio. Questa fase consuma e costa la cellula due molecole di ATP.
Tuttavia questa è indispensabile per guadagnare dell'ATP maggiore. Per questo motivo si chiama fase di investimento. Succede a questa la fase di recupero in cui dalla gliceraldide fosfato ricaviamo l'acido pirovico.
Ovviamente otteniamo due molecole di piruvato per ogni molecola di glucosio. Questa fase produce quattro molecole di ATP che sottratte alle due spese all'inizio nella fase di investimento ci danno già un guadagno di due molecole di ATP. In questa fase, inoltre, vengono liberati quattro elettroni.
Due per la trasformazione di ogni gliceraldosfato in piruvato. Questi elettroni assieme agli ioni H+ vengono utilizzati per trasformare il NAD+ in NAD H. NAD funge da trasportatore di elettroni, infatti ritrasformandosi in NAD più libererà due protoni e quattro elettroni.
Questo ogni due molecole di NAD. Vediamo innanzitutto cosa succede se la cellula è ossica, ovvero non c'è ossigeno a disposizione. Quello che avviene, come abbiamo detto, è la fermentazione.
Lo scopo della fermentazione è quello di utilizzare NADH e ritrasformarlo in NAD+. L'unica funzione della fermentazione è quella di rendere nuovamente disponibili delle molecole di NAD più per caricarsi gli elettroni che verranno liberati durante la fase di eh guadagno o di recupero della glicolisi. Questo quindi serve solo a permettere numerose glicolisi, ma non produce energia di per sé.
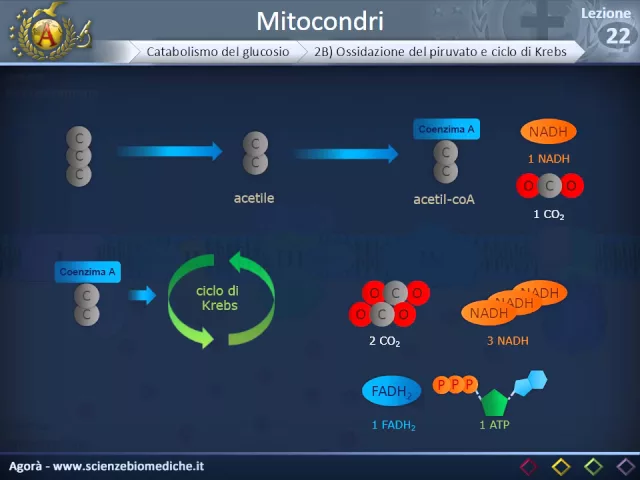
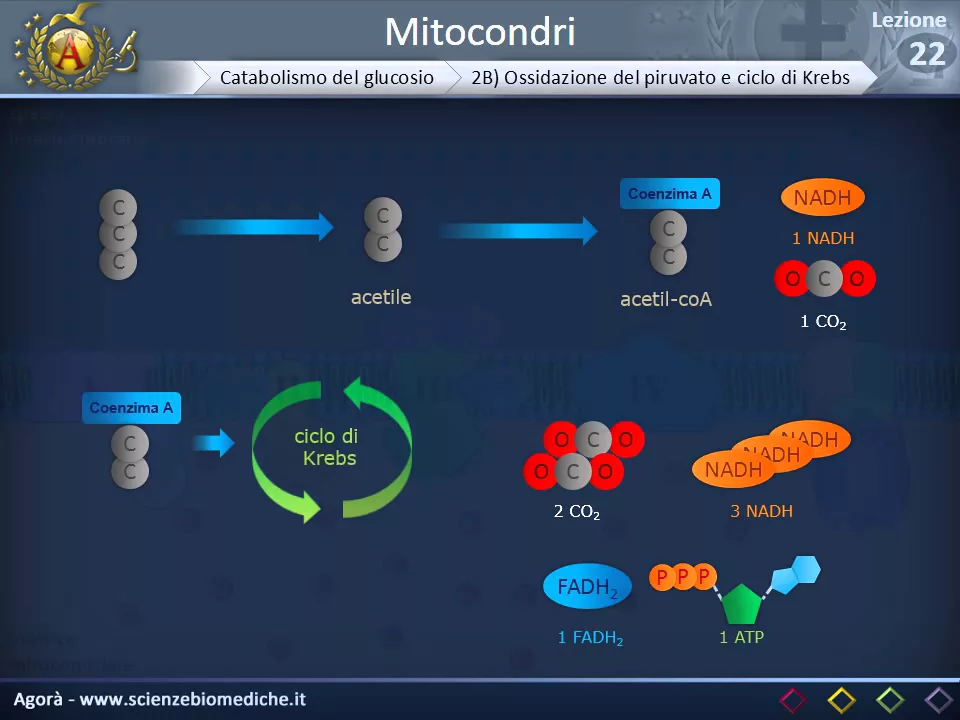
Nel caso della fermentazione, quindi, la produzione energetica è limitata solo all'ATP prodotto dalla glicolisi. Torniamo quindi alla respirazione cellulare, quella che avviene solo se c'è dell'ossigeno a disposizione. Il primo passaggio è trasformare il piruvato prodotto dalla glicolisi in acetile.
All'acetile viene poi aggiunto ilima A e produce quindi acetilenzima A nad H e una molecola di CO2. Il Quenzima può quindi entrare all'interno del ciclo di Krebs. In questo ciclo il quenzima va incontro a una serie di trasformazioni e questo determina la produzione di due molecole di CO2, tre molecole di NAD H, una molecola di FAD H2 che è un trasportatore di elettroni come il NADH ed una molecola di ATP.
Tutto questo però va moltiplicato per due in quanto questi sono i prodotti di un piruvato, ma la glicolisi produce due molecole di piruvato per ogni molecola di glucosio, quindi il tutto va raddoppiato. Vediamo ora l'ultima tappa, la fosforilazione ossidativa. Questa è la tappa in cui effettivamente vengono prodotte molecole di ATP in grandi quantità.
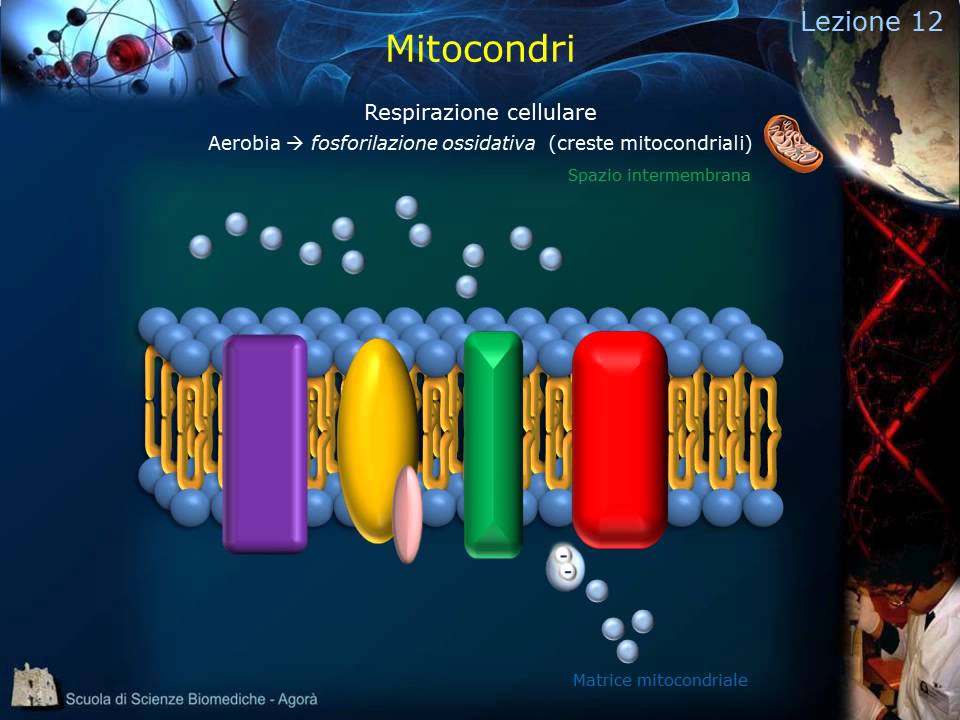
La fosforilazione ossidativa avviene sulla membrana interna. Questa possiede dei complessi trasportatori di membrana che una volta attivati pompano gli UNH+ concentrati nella matrice mitocondriale all'interno dello spazio intermembrana. Questo nuovo gradiente creato dai trasportatori sarà più utilizzato da una pompa di tipo F per creare ATP.
Vediamo come funziona. Il NAD H si riduce e cede due elettroni al primo complesso. Questo si attiva e pompa 4 H+ nello spazio intermembrana.
Questi due elettroni poi vengono passati sotto forma di atomo di idrogeno all'ubiquinone, una molecola altamente idrofoba che per questo motivo si sposta tra le code lipidiche della membrana interna. L'ubiquinone che si trova sul secondo complesso, quando riceve i due atomi di idrogeno, si sposta sul terzo complesso. A questo punto cede i due atomi d'idrogeno e li scompone fornendo un elettrone al citocromo C e un altro elettrone ad un wichinone scarico che si trovava sempre sul terzo complesso.
Questo attiva il trasportatore di membrana che pompa all'esterno altre quattro altri quattro ioni HP e l'ubiquinone torna a riposizionarsi sul secondo trasportatore. L'ubbiquinone può essere caricato anche in un altro modo, attraverso il FAD H2 che cede due ioni idrogeno, lo carica e questo nuovamente utilizza un elettrone per caricare un altro citocromo C. e un elettrone per caricare l'ubinone scarico presente sul terzo complesso.
Questo attiva nuovamente il terzo complesso, altri quattro ioni H+ vengono trasportati e l'ubquinone scarico si ricarica utilizzando due protoni e diventa lo biquinone H e può essere utilizzato per attivare nuovamente il complesso 3. Questo biquinone viene utilizzato solo per non sprecare ioni H+ ed elettroni. Notiamo inoltre che il secondo complesso non pompa mai un H+, ma è solo un mediatore per l'ubiquinone.
Vediamo ora come funziona il quarto complesso. Questo riceve due elettroni da due citocchomi C presenti sul terzo complesso. Una volta attivato, trasporta due ioni H+ nello sposo intermembrana e utilizza altri due ioni per legargli agli elettroni e formare due atomi di idrogeno.
Il quarto complesso lega inoltre una molecola di ossigeno, la scompone e lega ad ogni singolo atomo i due atomi di idrogeno, liberando acqua. Ovviamente per scomporre un'intera molecola di ossigeno sono necessari quattro elettroni in origine e quindi l'attivazione di quattro acitocromic. L'attività completa, quindi del quarto complesso, trasporta quattro ioni H+, due per ogni per ogni due citocromi C e produce due molecole di acqua partendo da una di ossigeno.
Alla fine il gradiente di ioni H+ che è stato generato dal primo, dal terzo e dal quarto complesso verrà sfruttato da una pompa di tipo F che produrrà una molecola di ATP ogni circa 3 ioni H+. Vediamo quindi di fare un piccolo riepilogo finale. Abbiamo detto che la glicolisi produce quattro molecole di ATP, però ne consuma due nella fase di investimento, quindi il guadagno finale della glicolisi e due molecole di ATP.
A questo punto, se non c'è ossigeno a disposizione, si avrà la fermentazione che non produce altre molecole di ATP, quindi il guadagno totale della fermentazione è due molecole di ATP per ogni molecola di glucosio. Se invece c'è l'ossigeno per avviare la respirazione cellulare, il guadagno sarà molto più alto. Il ciclo di Crebs produce una molecola di ATP per ogni acidirco A che entra.
Quindi avremo due molecole di ATP totali prodotte per una molecola di glucosio, perché ricordate che ogni molecola di glucosio viene scomposta in due molecole più semplici che diventeranno acidilcoenzima A. La fosforillazione ossidativa invece nel complesso consuma 28 ATP. Ricordiamo che la fosforelazione ossidativa utilizzerà tutti gli elettroni che sono stati caricati sui Nedh e sui fadh durante i vari passaggi fino al raggiungimento del ciclo di Crebs.
Con un semplice calcolo possiamo osservare che il catabolismo del glucosio attraverso la respirazione cellulare produce 32 molecole di ATP che sono nettamente più alte di quelle prodotte dalla fermentazione. Tuttavia il la respirazione è più lenta e richiede inoltre ossigeno e in condizioni di stress la cellula può non essere in grado di utilizzarla e deve accontentarsi della fermentazione. I mitocondri intervengono nella morte cellulare programmata causando l'apoptosi.
In realtà i mitocondri intervengono solo nella via di attivazione intrinseca e non in quella estrinseca. Ma per maggiore completezza parleremo di entrambe. La via estrinseca inizia la poptosi a partire da segnali esterni che solitamente sono tumor necrosis factor.
I principali sono TNF alfa, trail e faz ligando. Questi messaggi legano un recettore di membrana con dominio di morte. Quando tre domini di morte si accoppiano, attivano proteine che hanno un dominio di morte complementare, in questo caso la procaspasi 8.
La procaspasi 8 quando lega i tre domini di morte del recettore si attiva e diventa caspasi 8. La caspasi 8 a sua volta attiva la caspasi 3. La caspasi 3 poi darà l'inizio alla poptosi.
La ventrenza che invece è attivata da segnali interni. ATM è una proteina che riconosce i danni al DNA e quindi l'informazione della cellula. Se l'informazione è compromessa, la cellula non svolgerà più correttamente il suo compito e può diventare addirittura dannosa per l'intero organismo, come ad esempio nei tumori.
Quando ATM riconosce un danno, determina la produzione di P53. Questa proteina è importantissima e viene definita l'angelo custode del nostro genoma. P53, infatti, si occupa di riparare i danni al DNA richiamando anche altre proteine.
Se questi però non sono riparabili, P53 si accumula e un accumulo di P53 causa la poptosi, ovvero la morte, perché è preferibile che la cellula muoia, ma che si salvi l'intero organismo. L'accumulo di P53 determina la formazione di proteine canale come BX e BAC. Queste determinano l'apertura della membrana esterna del mitocondrio.
Queste verture formate da dimmerid di BX e BAC permettono la fuoriuscita del citocromo C dall'interno del mitocondrio. Quando il citocchomo C si si accumula nel citoplasma lega Apaf1. Apaffuno in presenza di del citocromo Cambia forma.
Il complesso ApaF1 e citocromo C forma una struttura a sette elementi, detto apoptosoma. Questa struttura lega la procaspasino e l'attiva rendendola caspasive. Questa richiama come la via estrinseca la Caspasi 3.
La Caspasi 3 rappresenta quindi sia per la via intrinseca che per la via strinseca l'ultimo passaggio che determina la poptosi. La Caspasi 3 ha quattro effetti: l'attivazione delle DNAI che spezzettano il DNA in frammenti regolari. Il collasso del citoplasma.
Il citoplasma, infatti, perde forma a causa della rottura del citoscheletro. La caspasi 3, inoltre, determina la frammentazione delle proteine che non devono essere più funzionanti. In ultimo, la Caspasi 3 termina la formazione di bolle apoptotiche.
La cellula si scompone in tante bolle pronte per essere facoitate in modo tale da non causare infiammazione. L'apoptosi però non avviene solo in condizioni patologiche quando la cellula è dannosa o non più funzionante, ma avviene in un gran numero di contesti fisiologici, ad esempio durante lo sviluppo embrionale. Nelle prime fasi abbiamo le mani palmate.
In seguito però le dita si separano grazie alla apoptosi generale di questo tessuto che le tiene unite. Senza la poptosi ci ritroveremmo ancora con le mani palmate. La poptosi, inoltre, serve a uccidere, a mandare a morte le cellule più vecchie e permettere che queste siano rimpiazzate da cellule più nuove.
L'apoptosi viene inoltre utilizzata per lo spegnimento della risposta immunitaria. Quando le cellule della risposta immunitaria non sono più necessarie perché il patogeno è stato sconfitto, queste cellule devono essere soppresse. La poptosi interviene oltre inoltre in meccanismi di atrofia dipendenti dagli ormoni, come ad esempio il caso della ghiandola mammaria all'interno dei maschi.
L'apoptosi è quindi una morte cellulare programmata. La cellula è consapevole di dover morire e quindi fa il possibile per non causare danni nella sua morte alle altre cellule. Essa infatti si scompone in vescicole e bolle apoptotiche per essere fagocitata senza danno per nessuno.
Scompone inoltre in modo autonomo, tutte le sue proteine e frammenta il DNA. L'apoptosi per questo è un processo attivo e consuma energia. Questa infatti differisce dalla necrosi che è una morte improvvisa, non programmata.
La necrosi può avvenire per un danno diretto alla cellula, come il calore o il taglio. Solitamente l'evento scatenante della necrosi è la frammentazione della membrana. Un'altra causa di necrosi può essere la carenza di ossigeno.
Se una cellula non riceve abbastanza ossigeno, non è più in grado di effettuare la respirazione cellulare e attraverso la semplice glicolisi seguita dalla fermentazione non produce abbastanza energia per rimpiazzare le sue membrane. A un certo punto la membrana esterna si rompe e questo è dannoso perché il contenuto interno della cellula con tutte le proteine ancora attive si riversa nell'ambiente extracellulare e interagisce con le altre con le altre cellule. Inoltre, anche le vescicole lisosomiali si rompono e gli enzimi lisosomiali si riversano all'esterno e questi sono dannosi.
La necrosi, quindi, è un processo passivo, non prevedibile e che causa danno per le altre cellule. Inoltre, mentre la poptosi è focalizzata su una singola cellula che riceve un messaggio dall'esterno dall'interno, la necrosi è un fenomeno che solitamente coinvolge un intero tessuto. Repilogando, quindi, i mitocondri intervengono sia nel metabolismo cellulare per la produzione di ATP e sia nella morte cellulare programmata o apoptosi.
Tuttavia, i mitocondri svolgono anche funzioni secondarie, però non meno importanti. Tra queste la proliferazione cellulare. Infatti la duplicazione dei mitocondri e il loro aumento dell'attività è fondamentale per fornire alla cellula l'energia necessaria per duplicarsi.
I mitocondri, inoltre, intervengono nella produzione di calore e i mitocondri si occupano anche della sintesi del colesterolo.