[Música] Hola cómo les va bueno La idea es la siguiente en esta en este caso la semana que viene nos vamos a meter de lleno en el estudio de esta vía metabólica una de las vías centrales del metabolismo intermedio que es la vía de los ácidos tricarboxílicos o la vía del ácido cítrico o la había conocida como el ciclo de krebs y dado que el acetilcoa es sustrato para esta vía nos parece importante hacer una primera aproximación de Cuál es uno de los posibles orígenes de esta molécula que es un punto interesante A considerar en
el metabolismo intermedio dada su conexión con carbohidratos con lípidos con aminoácidos con los aspectos energéticos Entonces nada vamos a hablar un poquitito de uno de los principales orígenes de esta muerte el acetín coa es un acetato activado por esta molécula que es la famosa coenzima este acetato activado termina siendo el punto central en la convergencia de vías catabólicas y divergencia hacia procesos de síntesis de otras biomoléculas el acetilco a es el producto del catabolismo de la glucosa el acetil codal es el producto de catabolismo de ciertos aminoácidos el acetílico a es el producto del catabolismo
de ciertos lípidos particularmente los ácidos grasos el acetílico a es sustrato para la síntesis de cuerpos cetónicos sustrato para la síntesis de ácidos grasos sustrato para la síntesis de colesterol que a su vez utilizamos para la síntesis de hormonas esteroides y el acetilco a es sustrato para el ciclo de krebs una vía que está netamente relacionada con la energía Pero además como vamos a ver está relacionado con la síntesis de otras biomoléculas por lo tanto en este simple esquema queda bien representado este papel tan central que va a tener el acetil coa como un punto
de convergencia y divergencia de los procesos catabólicos y anabólicos y dada esta situación y su conexión tan directa con el ciclo de krebs nos pareció interesante plantear la primera aproximación que es la aproximación desde el metabolismo de la glucosa Entonces el objetivo que tenemos para esta para esta charla es tratar de reconocer los puntos centrales de la formación de acetil Coat a partir del piruvato que proviene de el metabolismo de la glucosa encima encargada de este proceso se llama piruvato de hidrogenasa y en realidad como vamos a ver un ratito es un complejo multi enzimático
básicamente la pivoto es hidrogenasa cataliza la descarboxilación oxidativa del piruvato es decir que esta molécula de tres carbonos cuyo principal de origen es la glucosa pero que puede provenir también del lactato o de aminoácidos como la alanina pierde un grupo carboxilo y une una molécula de coenzima a para generar nuestro famoso acetilcoa el complejo piruvato es hidrogenasa es un complejo mitocondrial por lo tanto la síntesis de acetilcoa es mitocondrial Mientras que el piruvato es una molécula que se forma sea derivada de la glucosa sea derivada del lactato o sea derivada de aminoácidos a nivel cito
por lo tanto el primer punto de análisis es el ingreso de piruvato a la mitocondria esto ocurre a través de dos transportadores bastante ya reconocidos un transportador que encontramos en la membrana microcondrial externa y un transportador que encontramos en la membrana mitocondrial interna pero son los encargados de permitir el pasaje de piruvato del citosol hacia la matriz mitocondrial para que entonces la piruvato es hidrógenasa cataliza su descargo oxidación oxidativa el primer transportador de pibato en realidad es un transportador dependiente de voltaje y es un transportador para aniones es decir que no es un transportador que
sea específico para el piruvato sino que es un transportador para aniones y que el piruvato el ácido pirúvico aprovecha para poder meterse el segundo transportador sin Parece ser un transportador específico para el piru no se lo conoce como transportador mitocondrial para piruvato pueden llegar a encontrar que hay de tipo 1 de tipo 2 y es la conjunción de la acción de ambos transportadores lo que entonces permite el ingreso del piruvato citosólico independientemente del origen que tenga hacia la matriz mitocondrial para que Entonces ahora sí el complejo piruvato es hidrogenasa pueda catalizar esta reacción que están
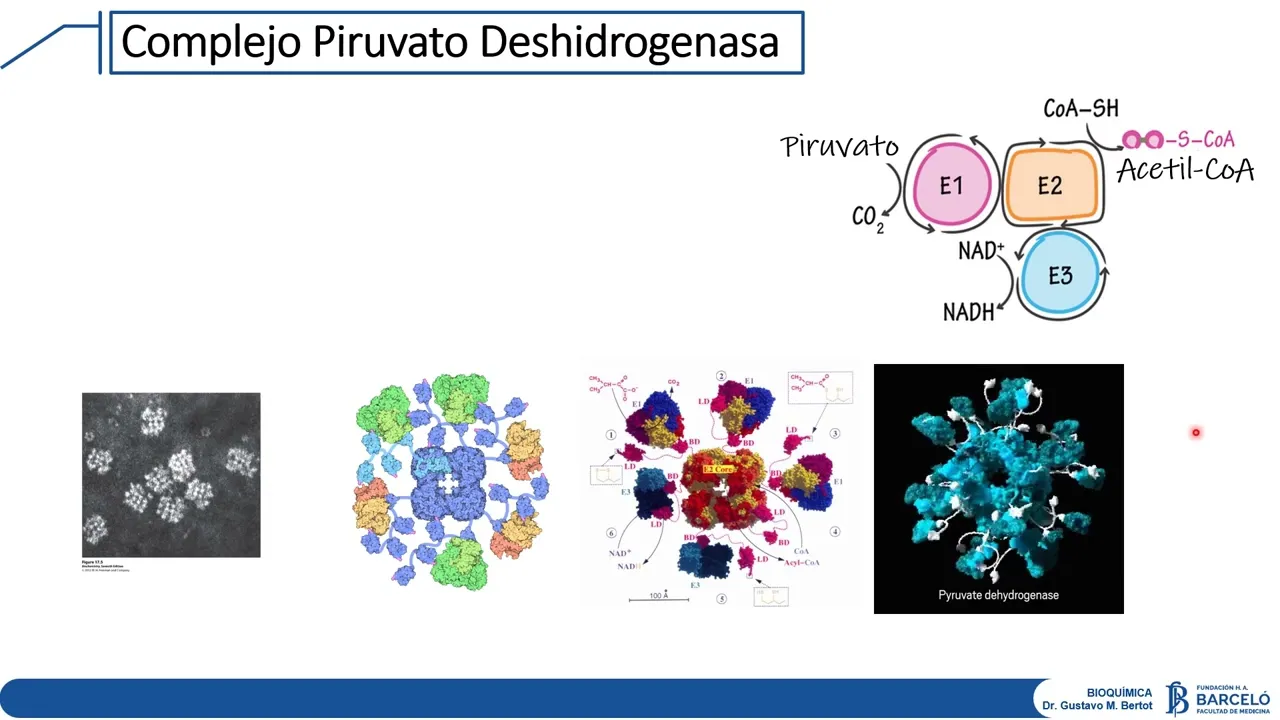
tan central en el metabolismo intermedio es un complejo en realidad multi enzimático está conformado por tres enzimas que participan del proceso catalítico que son una conectividad de carboxilasa la podemos denominar e1 una conectividad transaccetilasa la podemos denominar el 2 una deshidrogenasa propiamente dicha la podemos denominar e3 y tiene otras dos actividades enzimáticas que son una actividad quinasa y una actividad fosfatasa es decir que este complejo con cinco actividades enzimáticas me está mostrando tres de ellas que participan del proceso catalítico en sí mismo y dos de ellas que da la sensación que van a participar de
procesos de regulación dado que al tener una actividad quinasa y una actividad fosfatasa es muy posible que estén emparentados con procesos de fosforilación de fosforilación lo que es interesante es que la este acción coordinada de estas tres enzimas requieren la presencia de cinco cofactores enzimáticos vamos a vamos a nombrarlos y así volvemos a remarcar algunos aspectos sobre cofactores coenzimas etcétera tenemos como factor el pirofosfato de tiamina derivado de la vitamina B1 de la tiamina que está asociado a la primera actividad enzimática que es la descarboxilasa tenemos el ácido lipoico que no es una vitamina el
ácido lipoico es un derivado de un ácido graso de 8 carbonos del ácido octanoico tenemos la coenzima que se sintetiza utilizando ácido pantoténico tenemos Far asociado a la enzima A3 que ya hemos hablado algo es derivado vitamina b2 Y tenemos nada que si nos aparece explícito en la reacción que es derivada de la niacina uno podría preguntarse Por qué habiendo cinco cofactores necesarios para poder desarrollar la actividad de este complejo enzimático solamente en la reacción que yo escribo me aparece el nada La respuesta es porque los otros cuatro funcionan como grupos protéticos Es decir que
están unidos alguna de estas sus unidades enzimáticas razón por la cual no están apareciendo explícitamente en la reacción Recuerden que en algún momento comenzamos a ser diferencias entre lo que es una coenzima que tiene libertad de unirse a una enzima versus un grupo prostético que cumple todo su ciclo catalítico unido a la enzima esa es la razón por la cual solamente aparece en la reacción el nada dado que tiene la capacidad de poder unirse cuando el complejo lo necesita y dar como producto en este caso el nadache que se libera para digamos de última volver
a reoxidarse en cualquier otra enzima este con actividades hidrógenas o sea de tal manera que el digamos el funcionamiento coordinado de estas tres este unidades enzimáticas es lo que permite desarrollar la de carboxilación oxidativa que estamos hablando vamos a revisarlo solamente por revisarlo no para poder entender un poco Cómo es que se coordinan Y cómo es que estas enzimas digamos desarrollan su actividad nosotros digamos acá en la parte inferior tenemos el piruvato fíjense que el piruvato en principio ingresa a la enzima e1 Y en donde carboxila uniéndose al pirofofato de tiamina la encina de 2
es la que tiene el ácido lipoico este ácido lipoico les dije que era un ácido en realidad es un derivado de un ácido graso de 8 carbonos que a su vez se encuentra unido a un aminoácido lisignar son para lo cual este esquema es bastante este representativo fíjense que se forma como si fuera una especie de brazo que tiene la capacidad de poder establecer una conexión entre lo que debe suceder a nivel de la enzima de dos con lo que ha sucedido a nivel de la enzima e1 este ácido lipoico tiene sí fíjense tiene dos
grupos subfidrilos que son importantes para poder unir a este ahora acetato dado que acá hubo la descarboxilación y permitir la unión de por encima para que salga como producto nuestro hace cinco a el resultado de este momento enzimático Define que el ácido lipoico no ha quedado químicamente igual a como había arrancado la reacción y algo que cuando comentamos sobre las enzimas todos y digamos hicimos bastante hincapié era en el concepto que una enzima cumple todo su ciclo catalítico Pero al final permanecen alterada Esto vale también para los cofactores razón por la cual como a cada
un proceso redox este ácido lipoico tiene que volver a su estado original y la vuelta a su estado original es la necesidad de él la enzima e3 que aprovechando su actividad deshidrogenasa utiliza alfar y al nad como los como las moléculas que participan en la transferencia de electrones de tal manera que la oxidación del ácido lipoico rinde los electrones para generar los nada H como producto Entonces estos cinco cofactores enzimáticos están participando en forma directa en el proceso de la descarboxilación oxidativa y tienen la capacidad de poder digamos trabajar en forma coordinada para que el
complejo vuelva a su estado original digo que es un complejo enzimático importante fíjense una acá observamos Una Micro fotografía electrónica en donde observamos los complejos hidrogenasa que estarían conformados aproximadamente por un núcleo central de al menos 60 unidades E2 y una especie de cobertura con 30 sus unidades de uno seis sus unidades C3 más el complejo digamos la piruvatoquinasa y piruvato de hidrogenasa fosfatasa si uno mira una estructura actual de estos complejos podemos observar como en esta animación te da bien explícita la capacidad de interacción que tienen las subunidades E2 a través de esos brazos
que contiene el ácido lipoico para poder relacionarse con e1 y con e3 para producir la de carboxilación oxidativa habíamos visto un poco las características estructurales sobre todo la tan importante relación en requerimientos de cofactores enzimáticos de esta enzima que a su vez define un requerimiento de vitaminas vamos a analizar un poco los aspectos de control de esta enzima dado que parece ser un punto muy importante a nivel del metabolismo intermedio teniendo en cuenta que el acetilcoa es una molécula central para la síntesis de otras biomoléculas y para su ingreso al ciclo de krebs lo que
vamos a encontrar Es que la piruvato es hidrogenasa tiene una regulación alostérica muy importante por el por sus propios productos para además dada la presencia de la piruvato de hidrogenasaquinasa y de la priorato deshidrogenasa fosfatasa tiene su propio sistema de regulación covalente de tal manera que cuando la piruvato es hidrogenasa se encuentra desfosforilada se encuentra en su forma activa mientras que cuando se fosforila la enzima pasa a un estado inactivo sí Acá hay un concepto que es importante a tener en cuenta no estamos hablando de una cascada a través de proteínas GS alimentos y clases
en específico PK sino que estamos hablando que es la propia el propio complejo el que tiene en una de sus actividades enzimáticas la capacidad de autoforilarse o de desfosforilarse para así poder pasar de un estado inactivo A un estado activo vamos a analizar con un poquito más de detalle esta situación bien lo primero que dijimos es el control que establecen los propios productos de la reacción el acetilco a y el nadache de manera inhibitoria de tipo competitiva sobre la enzima esto significa que es la demanda de acetilcoa le va a ir jugando sobre la velocidad
de flujo de transformación de piruvato acetilcua Por supuesto Siempre y cuando haya piruvato disponible si la célula necesita acetilcuada para derivarlo a alguna vía en particular eso va a permitir un flujo continuo mientras que si las demandas de acetilco a disminuyen y aumenta su concentración este mitocondrial ese aumento inhibiendo la actividad está encima va a ir disminuyendo la velocidad de formación Y de esa manera se va a ir manteniendo una concentración equilibrada En los niveles de acetilco ámbitocondriales por el otro lado dijimos que teníamos La regulación covalente es decir su pasaje el estado fosforilado implica
la inactivación del complejo multienzimático sí los puntos centrales de fosforilación se encuentran en la enzima 1 y para eso es que tenían la piruvato y hidrogenasaquinasa y la piruvato es hidrogenasa fosfatasa como los autos sistemas de fosforilación de fosforilación que controlan a su vez la actividad de extensiva fíjense que este ciclo de fosforilación también se encuentra controlado para poder permitir una regulación coordinada con La regulación por modular por modulación este que generan los propios los productos de la de la reacción Quienes son los principales moduladores positivos de la actividad quinasa aparecen de nuevo el acetilco
a y el nadache es decir que los aumentos de acetílico a a nivel mitocondrial no sólo inhiben competitivamente a la enzima sino que a su vez estimulan a la piruvatos hidrogenasa quinasa para que pase el complejo a un estado fosforilado inactivo Y de esa manera la célula en ese momento en esa condición se está garantizando que no haya producción de acetilcuas no solamente se está garantizando que disminuya la velocidad sino que está llevando a muchas de esas enzimas a una situación de total inactividad es decir que en la medida que la célula tenga buenos niveles
energéticos disminuye la necesidad de Seguir generando lo que veremos sustrato del ciclo de krebs directamente asociado a la formación de energía por el otro lado el piruvato sí tiende a tratar de evitar que esta enzima sin active sino tratar de que sus flujos sea para la formación de acetilcuas lo que no podemos hacer en principio es trabajar el control de la actividad fosfatasa a través de una situación de tipo Espejo o de contraposición El este modulador más importante que activa a la piruvato descirogenasa fosfatasa es el calcio dado que una de sus isoenzimas une calcio
y magnesio por lo tanto lo que sí pueden es hacer Cualquier relación que tenga que ver con aumento de calcio Por ejemplo si estamos en el hepatocito en una condición posprandial en donde la insulina a través de su receptor está aumentando los niveles de calcio citosólicos y en consecuencia beber aumento de calcio mitocondrial ese aumento de calcio mitocondrial es un factor que va a activar a la piruvato deshidrogenasa fosfatasa para que entonces haya más disponibilidad de encima activa Porque seguramente la condición metabólica es para que se forme acetilguar y que tome destinos definidos como estudiaremos
más adelante entonces fíjense que acá hay una regulación coordinada no son compartimentos separados Dado que los mismos moduladores que regulan la actividad a través de lo que nosotros definimos como modulación de tipolostérica a pesar de que acá claramente es un fenómeno de emisión competitiva son los que controlan la actividad de la piruvato es hidrógenasa para llevar la enzima a un estado inactivo Me gustaría para ir cerrando un poco digamos esta charla y sobre todo por si lo llegan a leer es que tengan la siguiente consideración existen cuatro y su enzimas de piruvato de xilógenasaquinasa y
como Nosotros sabemos que la presencia de su enzimas y su expresión diferencial en tejidos es lo que nos va a permitir a nosotros entender realmente cómo va a funcionar esa reacción en el metabolismo celular vale la pena no para discriminar entre ellas sino reconocer que existen diferentes factores que pueden modular la actividad está encima en función de las necesidades particulares de un tejido de una célula por eso es que les traigo solamente estos ejemplos así como la insulina a través del calcio tiene la capacidad de poder activar la actividad fosfatasa Y de esa manera llevarla
encima un estado activo al mismo tiempo está inactivando a la piruvato y cirogénasequinasa para que esto no se transforme en un círculo vicioso es muy interesante Cómo es la respuesta de una célula alipoxia a la disminución en la presión parcial de oxígeno existe en la célula una proteína central que reconoce la fluctuaciones en la expresión parcial y oxígeno y en consecuencia disparas mecanismos moleculares para poder hacerle frente a esa situación uno de esos mecanismos moleculares es activar sí la piruvato es hidrogenasa quinasa de hecho en realidad más que activar lo que hace es induce una
de estas isoenzimas y en consecuencia lleva el metabolismo desde un plano potencialmente aeróbico dado que este acetil coágulo supone que ingresará el ciclo de krebs hacia un plano de inactivación de este sistema y disponer de este piruvato ahora no puede ser utilizado para la formación de acetilcuas seguramente hacia la formación del lactato el ayuno los ácidos grasos es decir la metabolización de ácidos grasos la presencia de corticoides el ácido retinoico son factores que inducen la expresión de esta esquinazas y llevan el metabolismo nuevamente para que no haya flujo de piruvato acetilcóat y en tal caso
la célula encuentre nuevos desvíos metabólicos de ninguna manera estamos diciendo que obligadamente no haya acetilcoa sino que lo que estamos comentando es que es esta origen de acetilco a partir del piruvato posiblemente a partir del metabolismo de la glucosa lo que en tal caso en esta condición metabólica estaría comprometiéndose Me parece que esta es una primera aproximación que bastante más adelante Bueno nos metamos más el metabolismo de glúcidos y de lípidos va a empezar a tomar bastante bastante relevancia Pero bueno vale la pena hacer esta primer consideración En conclusión estamos hablando de un complejo enzimático
muy central en el metabolismo intermedio es la el encargado de generar Quizás El acetilco a través de una de las vías principales que es vía metabolismo de la glucosa un complejo muy enzimático mitocondrial con una complejidad estructural interesante la conjunción de cinco actividades enzimáticas tres para poder producir el proceso catalítica y dos que participan de su control un control que es muy fino porque tiene que establecer de forma adecuada el equilibrio entre la oferta la oferta de piruvato y la demanda de acetilco a de tal manera que en forma concertada nos permite entender esa regulación
por inhibición competitiva y regulación de tipo ecovalente un complejo multi enzimático que tiene una un requerimiento interesante en vitaminas dado que tiene cinco cofactores enzimáticos cuatro de los cuales este son digamos están relacionados con vitaminas pirofosfato de tiamina tenemos la coenzima a tenemos el Far Y tenemos el nad y que trabajan de manera coordinada para poder cumplir con ese famoso postulado que una enzima y sus grupos prostáticos deben permanecer en altira inalterados al final de la reacción así que bueno esta primera aproximación sobre uno de los orígenes del acetílico a Y entonces ahora sí por
lo menos ya tenían una referencia de dónde va a venir el acetilco a como una molécula tan central de metabolismo intermedio la próxima nos adentraremos en uno de sus destinos principales el ciclo de krebs gente Gracias por escuchar nos vemos cuídense
![Upbeat Lofi - Deep Focus & Energy for Work [R&B, Neo Soul, Lofi Hiphop]](https://img.youtube.com/vi/THh4fT0O7IY/maxresdefault.jpg)