okay so welcome to this next video in the playlist on cancer in this video what we're going to talk about is colorectal carcinoma and in particular we're going to look at a model for what happens in colorectal carcinoma so an actual model that outlines exactly what happens now of course um you have to remember all science is just models for the real world and you might and this model that i'm going to show you the vogelstein model is very very simple and given the complexity that could occur in cancer uh you might think that it's
over simple what i will say in the defense of the vogelstein model is that it's a very simple model nice and easy to understand and it actually is remarkably good at making predictions which if you then do experiments and um experimental inquiry basically the experimental data generally agrees with this model so it's very very good it's a good model basically and it's very simple it's what's nice about it okay so um let's begin then that's the way i'm going to structure this by the way is we're just going to work through the vogelstein model and
we're going to see the mutations that occur at what point and while when we look at each of the mutations i will um briefly remind you of uh the pathways that each one is involved in and how that's going to lead to the effect that i claim it has okay and i want hopefully that this um discussion of the vogelstein model of colorectal carcinoma will basically um also help you understand the the basic principles of all cancer because a lot of the basic principles that we're going to see here such as it being a multi-step
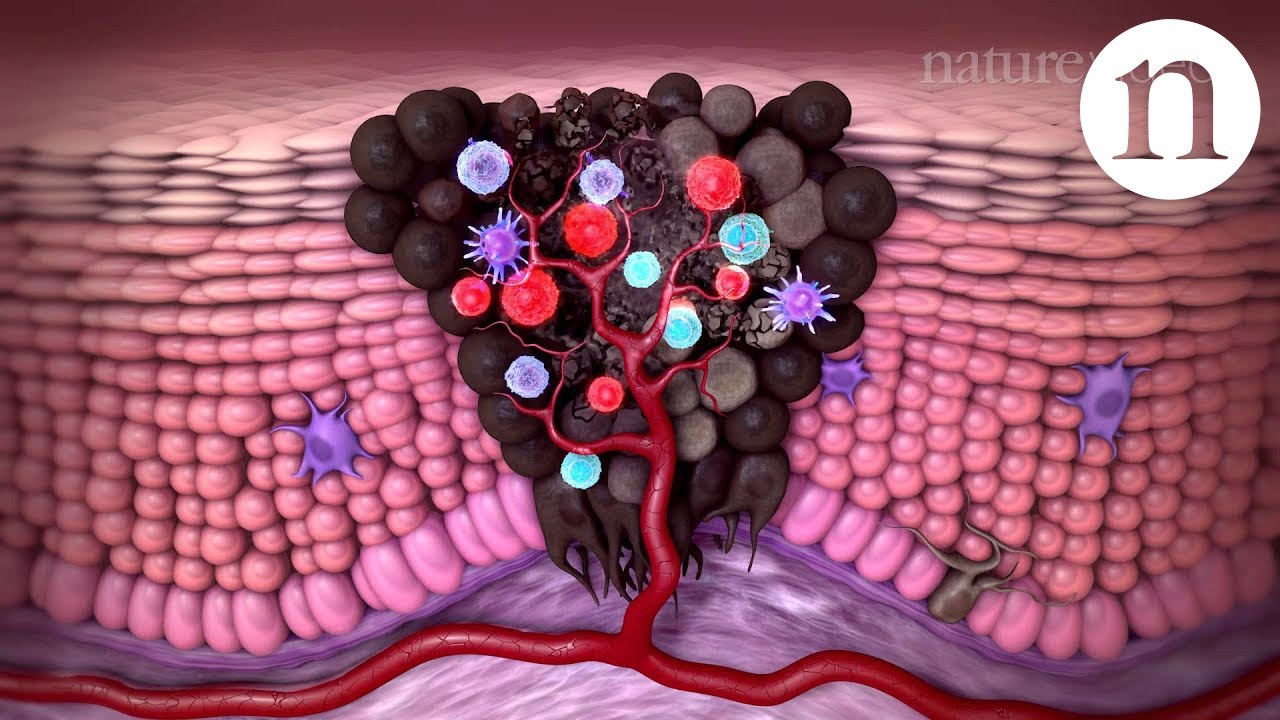
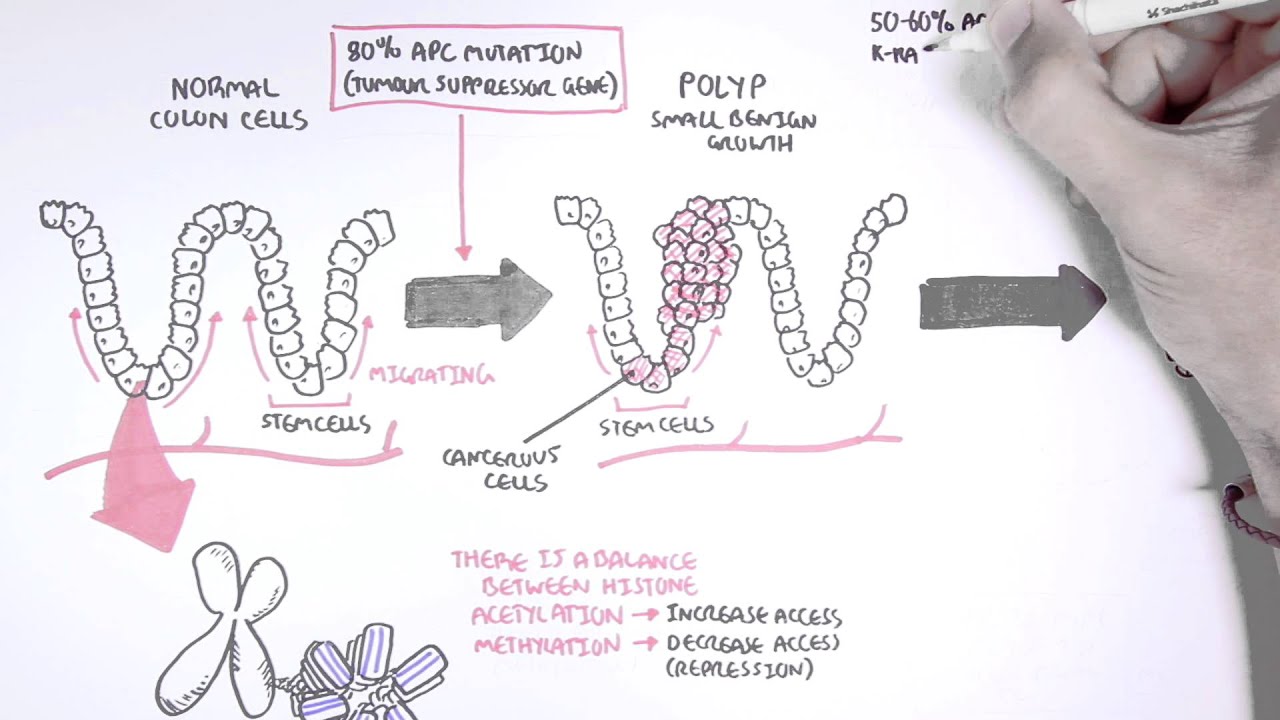
process it's not one mutation and then it's cancer it's a multi-stage process and the concept of intra-tumor heterogeneity i hope that those are going to become apparent in this video okay so let's begin let's say here we have the normal epithelium of the colon then okay so let's say these are the um villi let me just okay um and let's say that um well actually they're they're not vivi in the colon they're colonic crypts rather rather than villi so instead of um instead of the being imaginations out of the endothelium epithelium instead there's invaginations in
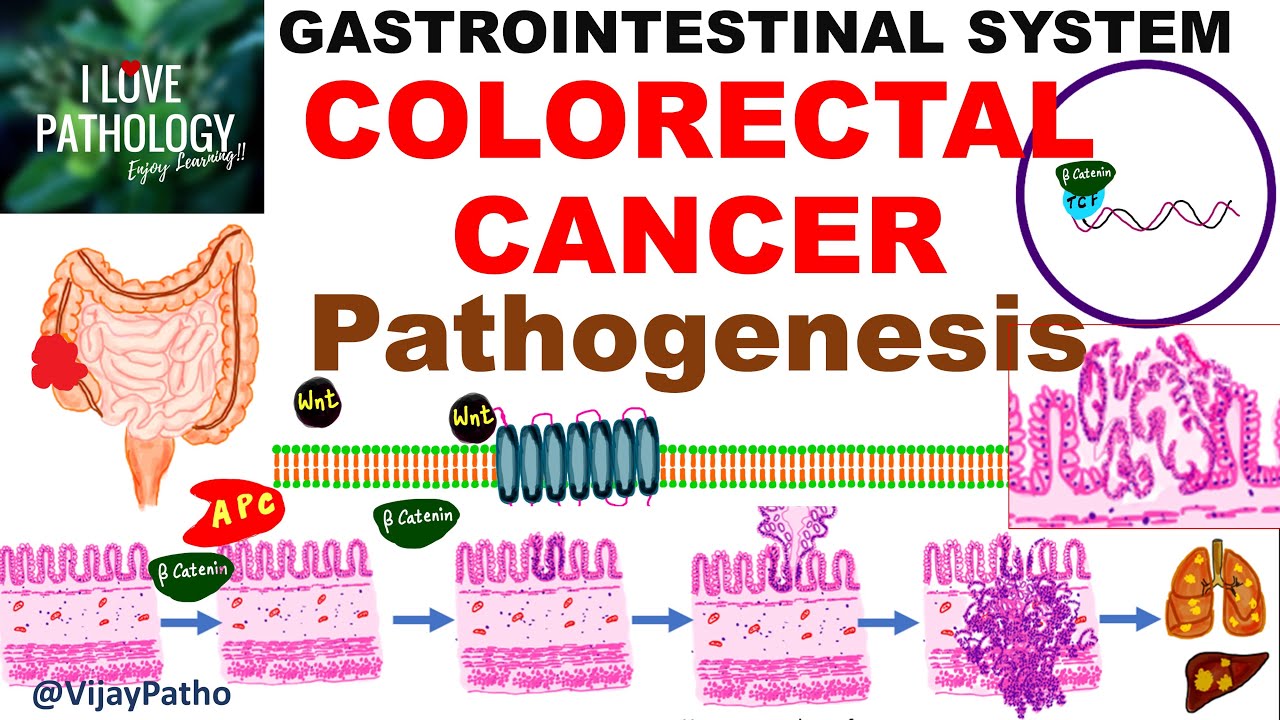
and these are the colonic crypts okay so let's say this is the normal colonic epithelium here okay and these are epithelial cells here let's divide some of them up and we want to see how you go from having normal epithelial cells to having a colorectal carcinoma or colon cancer okay right and this is the vogelstein model for how this happens i'll just put that the vogelstein model right okay so let's look at the first thing which happens in the vocal steam model of colorectal carcinoma so basically what has to happen is in a single cell
from this colonic epithelium so this is the colonic epithelium what has to happen is you have to get a loss of function in um you have to get a lot of function in the apc gene or the adenomatous polyphase coli gene okay so um let's have a brief reminder of the wind beta catenin pathway so that we can understand why a lot of function of the apc protein will lead to overgrowth of this cell that's going to get this mutation okay so basically uh wind is a growth uh inducing molecule which binds to receptors on
the surface of cells and these receptors consist of two actual proteins that make them up okay so one is a protein with a single membrane spanning domain like so and the other is a protein with a seven membrane a transplanting domain okay so this larger one with the seven membrane spanning alpha helices this is the frizzled receptor so this is frizzled and uh this other receptor that also um combines with friso receptor in order to make this receptor for wind this receptor here is the lrp4 slash 5 so either you can use the protein lrp4
or you can use the protein lrp5 okay so together lrp4 or 5 uh with this frizzled receptor which is often denoted just by f z so if you see people referring to f set that just means friso receptor okay these make up the receptors in the membrane of cells for the wind ligand so frizzled is a seven transmembrane receptor and finally let's just color in the ligand itself this wind here okay so this is wnt or wind right so when wind binds to the winch receptor which is made up of this frizzled receptor along with
this lrp4 5 protein what happens is it the frizzled receptor becomes active and it recruits another protein in here basically so let me draw that another protein is going to be recruited in here which unfortunately i've drawn my arrow right through where i wanted to draw the protein and we'll draw this protein in orange okay so this protein also has a silly name it's known as disheveled so this is disheveled disheveled okay and um disheveled is often uh also uh referred to as dsh for short so if you see people referring to dsh that's what
um it means it means disheveled now basically the wind signaling molecule is going to lead to the cell receiving a pro a pro-division stimulus so it's going to promote the division of the cell and in order to do that what you need to do is raise levels of beta-catenin in the cytoplasm of the cell so by activating this structure here this frizzled receptor well this winch receptor um and then uh the dischival disheveled protein you somehow need to lead to a rise in beta catenin so in order to understand how that happens we need to
look at the usual mechanisms which prevent beta catenin from elevating in the cytoplasm so basically beta-catenium is what's known as a transcriptional co-activator and we'll discuss further later what that means and usually to stop beta-catenin levels going up in the cytoplasm there is what is known as a beta-catenin destruction complex so let's have this big scary uh structure here okay so this is a beta catenin destruction complex so i will have this in pink okay and basically what the b-techteen and destruction complex is going to do is it's going to add a phosphate group onto
the beta-catenin molecule over here and that will target the b-ticketing molecule for ubiquitination so this is the beta catenin destruction complex and it's a complex of loads of proteins and we're going to see which proteins in a moment so this is the beta catenine destruction complex okay right so as i told you what it does to beta-catenin is it phosphorylates beta-catenine so if this is beta-catenine it's going to stick a phosphate group onto beta-catenin like so so here's a phosphate group on the side of b-ticket even and i'm going to color in b to continue
we'll have it in yellow okay so beta-carotene in yellow here has now got a phosphate group stuck on the side of it and once it gets a phosphate group stuck on the side of it uh that acts as a group onto which a ubiquitin group can then bind so this marks it basically for ubiquitination so the beta catenin destruction complex itself does not add the ubiquitin group onto beta-catenin instead what it does is it adds this phosphate group onto um beta-catenium which can then uh have a ubiquitin group added on to that so here is
our ubiquitin group being added on to the phosphate group okay right so i will color in the ubiquitin group here okay so in green right um so um things which get ubiquitin added onto them basically get targeted for destruction by the proteosome so the proteasome recognizes this ubiquitin molecule and anything that has ubiquitin uh attached to it such as now beta catenin here in yellow is going to get destroyed by the proteasome so the proteasome is basically a large tunnel that exists in um exists in the cytoplasm of cells and proteins go in one way
and amino acids come out the other side so it's going to destroy this beta-catenine basically so what will happen is in a usual cell which is not dividing beta catinium will be made and then the b-ticketing destruction complex will be active and it will phosphorylate beta-catenine instantly uh well pretty quickly and uh then that beta catenium will get a ubiquitin group added onto it uh which is the process known as ubiquitination okay so this is ubiquitin nation and then once it's got the ubiquitin group added onto it it will then be destroyed by the proteasome
so it won't ever do anything within the cell now this beta catenan destruction complex thus is very important for stopping beta-catenin from doing anything so let's now look at the proteins which make up the betacatene and destruction complex so there are five proteins which make this up uh and we'll talk about all five of them we'll give all their names anyway so one is something known as glycogen synthase kinase free or gsk free shorts this stands for glycogen synthase kinase free okay so you would think that that was involved in metabolism and indeed um glycogen
synthase kinase enzymes are involved in metabolism but that also this specific type this glycogen synthase kinase-free is also involved in this beta-catenin destruction complex so there's an example of how proteins can perform multiple roles within a cell then we have something known as axin which is another protein then we have apc which is going to be the important one that we're discussing uh which stands for adenomatous polypsis coli so adenomatus polypsis coli polyposis coli okay then we have a casein kinase one alpha and sorry that should have an s in there as well sorry so
it's csk one alpha standing for casein uh kinase one alpha okay so the k the c and the s come from the casein and the k then comes from the kinase and then finally this one here is something known as pp2a which stands for protein phosphatase 2a so this is protein phosphatase 2a right um so phosphates 2a so these five components make up this beta catenin destruction complex and basically what's going to happen is when wind uh binds to its uh receptor on the surface of the cell uh it's going to activate this dishevelled protein
and the dishevelled protein will then bind to these beta-catenin destruction complexes and inactivate them so this disheveled protein is going to inactivate the beta-carotene and destruction complex so if the beta-catenin destruction complex becomes inactive then the beta catenine isn't going to be phosphorylated then it's not going to be ubiquitinated and it's not going to be destroyed so beta catenium will start to build up in the cytoplasm of the cell therefore when this beta-carotene destruction complex complex goes down while the activity of it goes down because it's being activate inactivated by the frizzle receptor and we'll
continue this discussion in the next video