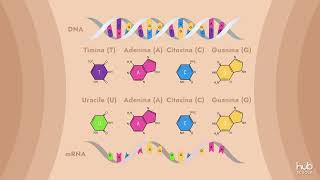
biologia lezione 13 in questa lezione finiremo di trattare l'argomento iniziato nella lezione precedente cioè l'rna la trascrizione e la traduzione con particolare riferimento ai processi di trascrizione e traduzione vediamo Innanzitutto il processo chiamato trascrizione la trascrizione quindi è il processo mediante cui il DNA Viene aperto viene trascritto in RNA dopodiché viene richiuso il trascritto sarà non non congruente allo stampo originale bensì complementare la complementarietà rispetto all stessa legge della complementarietà dell'appagamento delle due eliche del DNA con L'unica differenza che al posto della timina l'rna presenta la base azotata uracile questo processo viene portato avanti da
un enzima chiamato RNA polimerasi che sintetizza l'rna in direzione 5 Prim 3 Prim ovviamente il processo presenta delle caratteristiche diverse in procarioti ed eucarioti cominciamo a parlare dei nei procarioti la iscrizione presenta una fase di inizio una fase di allungamento e una fase di rilascio la fase di inizio è la prima fase nella fase di inizio il DNA Viene aperto ogni Gene da trascrivere presenta esoni ed introni Ma soprattutto è molto importante notare che nella parte subito prima del Gene da trascrivere è presente una piccola regione chiamata promoter noteremo che è composto da diverse sequenze
una posta a-35 e l'altra a-10 dove il numero negativo indica il numero di basi prima dell'inizio del Gene effettivo da trascrivere queste due sequenze servono per l'attacco della RNA polimerasi in particolare per l'attacco del fattore Sigma un cofattore che serve appunto all' RNA polimerasi per legare il promotore In particolare le subunità Sigma 2 e Sigma 4 legano le due parti speciali del promotore a-10 e-35 tra le due è presente un segmento di 19 nucleotidi La sequenza a-35 è composta da 6 nucleotidi La sequenza a-10 è composta invece da altri se nucleotidi il fattore Sigma è
necessario per la trascrizione Perché serve per individuare il sito di inizio della trascrizione serve per consentire il legame iniziale con il DNA e oltretutto Modula la velocità di lettura e scrittura vediamo un po' meglio come sono fatti questi promotori Innanzitutto la sequenza che si trova a-10 è formata da tre basi a g e t per esempio i promoter forti sono quei promoter che hanno una tripletta all'interno della zona che sta a 9 che è molto simile alla prima tripletta del Gene da trascrivere i promoter forti comportano un'alta velocità di scrittura della copia di RNA promoter
deboli invece presentano una disomogeneità tra la zona del è la prima parte del Gene da trascrivere questi promoter sono quindi deboli Nel senso che non inducono una trascrizione veloce quindi il gene trascritto in modo più lento si esprimerà di meno diamo ora un'occhiata al vero e proprio processo Innanzitutto l'rna polimerasi scorre fino a trovare il promotore Dopodiché apre una bolla di 35 nucleotidi all'incirca chiamata bolla di trascrizione lega il DNA in maniera quasi stabile Dopodiché avviene all'inizio vero e proprio inizia ad agganciare dei ribonucleotidi inizia quindi la fase di allungamento nella fase di allungamento il
DNA viene trascritto in direzione 5 primo primo il processo è sempre lo stesso vengono aggiunti i nucleotidi usando come stampo Il DNA è interessante Notare le subunità rudder e zipper che svolgono dei ruoli importanti la radder è molto utile nella nell'aggiudicazione è di 50 nucleotidi al secondo l'allungamento quindi prevede che il filamento di RNA si formi appaiato al filamento di DNA segue infine la fase del termine del rilascio ci sono due modi per il termine e rilascio due modalità completamente diverse La prima è quella dei degli RNA cosiddetti Ro indipendenti in questo caso il filamento
stampo di DNA presenta una grande quantità di basi GC e alla fine presenta molte adenine viene quindi formata una forcella che si Ripiega a causa dell'alto contenuto in g e c e questa indebolisce il legame della polimerasi con il DNA il colpo di grazia viene dato dalle ultime basi poiché il legame tra adenina e uracila è molto debole quindi l'rna polimerasi si stacca da sola senza bisogno di interventi esterni l'rna fa anche da terminatore In altri casi invece il terminatore è costituito da un enzima vero e proprio chiamato R per i geni che non hanno
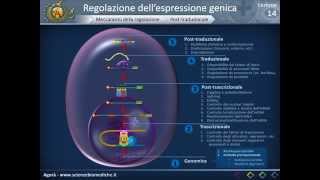
il terminatore R indipendente deve intervenire appunto la proteina R che una atpa sia adanello il cui compito è sfilare l'rna dalla polimerasi e staccare l'enzima dal DNA in questa immagine uno schema riassuntivo del concetto appena spiegato esistono ovviamente delle differenze tra la trascrizione eucariote e la trascrizione procariote nei procarioti abbiamo visto che c'è un solo tipo di RNA polimerasi mentre agli eucarioti ne presentano ben tre nei procarioti c'è un solo c Negli eucarioti ce ne sono molti nei procarioti non abbiamo elementi di controllo come gli attivatori repressori che invece sono presenti Negli eucarioti nei procarioti
c'è un solo promotore Negli eucarioti ci sono molti promotori proprio perché esistono molte ren polimerasi infine i procarioti non hanno elementi regolatori a lunga distanza Cosè che accade per gli eucarioti cominciamo a parare del della prima differenza gli eucarioti hanno ben tre RNA polimerasi l'rna polimerasi 1 sintetizza gli RNA ribosomiali 28 18 e 5.8s RNA polimerasi 2 si occupa soprattutto dell' RNA messaggero ma anche di alcuni piccoli RNA nucleari nucleoli e i micro RNA l' RNA polimerasi tre infine tRNA l'rna 5S e alcuni particolari tipi di piccoli RNA nucleari e citoplasmatici Esistono poi altre forme
più rare che sono l' RNA polimerasi 4 e 5 che sintez sintetizzano i crna nelle piante gli RNA polimerasi mitocondriale e dei cloroplasti che si trovano appunto all'interno di questi organolithium era riguardante i fattori di trascrizione Negli eucarioti c'è un gran numero di fattori di trascrizione Innanzitutto tutti i promotori o quasi tutti i promotori eucariotici presentano una sequenza chiamata TAT box il tatabox viene identificato e legato dal transcription Factor conosciuto come TF 2D TF 2D è fatto di due subunità la tbp che è il l tata binding protein e il TF questo insieme di fattori
richiama il TF 2A e il TF2 B che stabilizzano il legame con il DNA A questo punto viene richiamata la RNA polimerasi con il fattore TF2 F si aggiunge a questo complesso poi TF2 h TF2 J e TF2 Questo è il complesso di pre inizio il macchinario della trascrizione che la maggior parte dei fattori viene abbandonata e rimangono solo TF2 d e TF2 hf2 h attività elicas Quindi inizia a sciogliere i legami delle due eliche di DNA e la RNA polimerasi può iniziare la trascrizione vediamo Ora gli attivatori repressori Negli eucarioti sono presenti un gran
numero di elementi che si trovano a piccola distanza dal Gene da trascrivere e sono chiamati enhancer e silencer Questi pezzi di DNA hanno la funzione di aumentare o ridurre la trascrizione di un determinato Gene e lo fanno agendo sui fattori di trascrizione attraverso delle molecole chiamate attivatori gli attivatori non legano direttamente i fattori di trascrizione ma lo fanno attraverso un complesso mediatore abbiamo Inoltre detto che Negli eucarioti ci sono più promotori il complesso promotore eucariote è diverso che si se si parla di polimerasi 1 polimerasi 2 o polimerasi 3 per l' RNA polimerasi 1 abbiamo
il seguente complesso c'è il gene che ricordiamo essere quello dell' rrna 28s 18 s e 5.8s c'è il gruppo promotore Core che si trova tra - 45 e + 20 basi quindi può entrare anche all'interno del Gene il core viene legato dal TF 2D che in questo caso presenta tre subunità TF poi a 80-17 basi c'è l'upstream control Element che lega un fattore chiamato ubf che sta per upstream binding Factor e il meccanismo della trascrizione si esplica attraverso il piegamento del DNA che fa venire in contatto TF 2D e ubf e l'insieme di questi due
fattori richiama l' RNA polimerasi che inizia la trascrizione dei geni ribosomiali per la RNA polimerasi 2 c'è il gene che codifica solitamente per mrna ma anche per alcuni piccoli RNA come gli SN gli Snow e i micro RNA poi Ci sono alcuni elementi che fanno da Core che si trovano Tra - 35 e + 30 basi poi ci sono alcuni elementi regolatori prossimali che sono attivatori o repressori e si trovano tra 50 e-200 basi e infine ci sono enhancers e silencers più altre sequenze che si possono trovare oltre le meno 1000 basi Questi sono gli
elementi regolatori di stali ognuno lega un attivatore o un repressore l'rna polimerasi 3 Poi ha quattro tipi di promotori diversi il primo per l' RNA 5S è un promotore interno doppio il box a e il box C il secondo per i tRNA è sempre un promotore interno ma presente il box a e il box B per gli snrna abbiamo un promotore completamente esterno che comprende del tatabox Locked e il pse Infine per l'rna 7sl la cui utilità vedremo in seguito abbiamo un promotore misto che presenta i box A e B interni più il tata e
il TF adesso parliamo dell'ultima fase del dogma centrale della biologia cioè la traduzione la traduzione è il processo mediante cui dal RNA messaggero transfer e ribosomiale si ottengono delle proteine fanno parte di questo processo anche alcuni enzimi come l' Minino cilti RNA sintetasi i fattori di traduzione attenzione e gli chaperon e le Fold Asi la prima fase è con conosciuta come attivazione degli amminoacidi l'attivazione degli amminoacidi consiste nella nel legame a un semplice amminoacido di una certa quantità di energia fornita dal GTP l' Minin cil tRNA sintetasi lega sia il GTP che l'aminoacido dopo idrolizza
due fosfati del GTP e lega il gmp forma quindi un intermedio chiamato aminoacil gmp a questo punto interviene l'rna transfer che si lega in un terzo sito attivo e l'enzima a questo punto può collegare l'aminoacido al tRNA sleg il gmp che cede quindi la sua ultima parte di energia immagazzinando nel legame tra l'aminoacido e il tRNA a questo punto avviene il processo di proof Reading cioè avviene il controllo che l'aminoacido legato al tRNA sia quello giusto C'è un errore ogni 40.000 accoppiamenti subito dopo abbiamo la fase di inizio l'inizio avviene quando il ribosoma è ancora
separato il ribosoma presenta una subunità maggiore che prima D inizio inibita dall' if6 e una subunità minore che è legata sempre ai fattori if1 e i3 Intanto il tRNA con l'aminoacido legato è legato ai due fattori if2 ogni if2 è portatore di un pacchetto di energia sotto forma di GTP l'am Mino acil tRNA lega la subunità minore Questo si chiama complesso di pre inizio il complesso di pre inizio a questo punto Individua l'rna Come può individuarlo Perché l'rna lega altri fattori di inizio chiamati f4a i F4 a e g f4g i f4g è legato alla
Poli a dell' RNA messaggero la coda di lunghe ad if f4a a questo punto lega il CAP la set metilguanosina per controllare che l'aggancio sia ottimale in seguito il ribosoma Lega questi fattori formando il complesso di inizio vero e proprio e scorre fino a trovare la tripletta Aug che segnala l'inizio della parte da trascrivere a questo punto abbiamo il rilascio di un if2 sotto forma di idrolisi del GTP che permette la distruzione del fattore if6 cosa che libera la subunità maggiore del ribosoma che andrà a legarsi alla subunità minore la subunità maggiore presenta tre tasche
una è chiamata e la seconda è chiamata P La terza è chiamata a il primo tRNA si lega nel sito P il sito a è detto aminoacidico In quanto ogni nuovo amminoacido che entrerà nella proteina si legherà al sito a il sito P è detto peptidico perché ogni caena peptidica alloggerà sempre nel sito P il sito 3 È detto anche e o Exit vuol dire uscita In quanto ogni tRNA che si troverà nel sito Exit verrà slegato dall' RNA messaggero e mandato via vediamo ora la fase di allungamento nella fase di allungamento abbiamo l'intervento di
altri aminoacil tRNA che portano insieme a loro un ef1 elongation Factor One lef1 è portatore di una quantità di energia sotto forma di GTP l'aminoacido si lega al sito a poi viene spesa l'energia del GTP e l'aminoacido presente sul primo tRNA si sposta sull RNA appena entrato si inizia quindi a formare un peptide a punto interviene l'elongazione ribosom e sfruttando l'energia dell'idrolisi spinge il ribosoma spostando il peptide nel sito peptidico e il tRNA vuoto nel sito di uscita Interviene a questo punto un altro aminoacil tRNA con il fattore di allungamento 1 portante anch'egli una certa
quantità di ener che viene spesa per far avvenire il legame peptidico tra i primi due aminoacidi e l'aminoacido appena portato il termine e rilascio è l'ultima fase della trascrizione in senso stretto arrivati nella fase terminale ef2 spinge il ribosoma sfruttando come energia l'idrolisi del GTP e lo porta sull'ultima tripletta che è una tripletta non senso a questo punto Rimane solamente il tRNA del sito peptidil perché quello del sito Exit viene rimosso e nel sito amminoacidico si lega un tRNA vuoto perché parliamo di una tripletta non senso a questo punto il peptide si sposta sul tRNA
vuoto ma questo spostamento catalizza una rottura idrolitica del legame per cui si libera una molecola d'acqua e il peptide si slega dal tRNA diventando indipendente l'ultimo procedimento è il taglio della metionina il primo aminoacido iina infatti fa solo da segnale serve solo per iniziare la traduzione in quanto la tripletta di inizio è sempre la tripletta Aug che codifica per la metionina terminata quindi la traduzione la metionina deve essere rimossa Infine c'è la fase delle modifiche post traduzionali queste modifiche sono operate da alcuni organ particolari abbiamo l'aggiunta di glucidi chiamata glicosilazione l'aggiunta di lipidi che induce
un ripiegamento molto particolare della proteina abbiamo anche la formazione di ponti di solfuro tra due estremità molto lontane della stessa proteina abbiamo poi la fosforilazione molto important ante per attivare o deat alcune proteine e abbiamo anche semplici reazioni di taglio proteolitico le modifiche post traduzionali servono perché la proteina da semplice collana di perle cioè da semplice insieme di aminoacidi diventi una proteina perfettamente ripiegata con strura con struttura 3D precisa in quanto la funzione di tutte le proteine è legata alla loro struttura tridimensionale detta anche struttura terziaria il tempo di ripiegamento cioè di acquisizione della struttura
terziaria genera ha generato uno dei più conosciuti paradossi della biologia Il paradosso di Levin se Infatti si considera che il numero di conformazioni possibili per una una proteina di 100 aminoacidi e si suppone che ogni amminoacido possa esplorare al massimo tre conformazioni si ottiene che questo numero è pari a 10 all 87 se una proteina dovesse esplorare casualmente tutte queste combinazioni per ripiegarsi impiegherebbe 20 miliardi di anni il paradosso di levinthal vuole esporre come il ripiegamento delle proteine non sia casuale ma avvenga secondo delle leggi ben precise il biologo dawkins ipotizzò infatti che non è
che una proteina deve provare 10 all 87 combinazioni semplicemente ogni volta che un amminoacido va a posto cioè occupa la conformazione corretta quello si blocca e non può più tornare in una posizione scorretta secondo dawkins questo avviene perché la struttura primaria delle proteine cioè la sequenza di aminoacidi è fatta già in modo da avere intrinsecamente la capacità di ripiegarsi in maniera tridimensionale molte delle modifiche post traduzionali che riguardano i ripiegamento vengono operate dalle Fold Asie e dagli chaperon dette anche Head shock proteins gli chaperon sono delle proteine molto particolari vengono prodotte quando l'organismo viene sottoposto
a shock di calore perché il calore denatura le proteine Allora la produzione di chaperon serve a rinatura durante la febbre ci sono anche altre condizioni come gli stati patologici o alcuni fattori di crescita che inducono la formazione di chaperon Esistono gli chaperon veri e propri detti anche chaperon propriamente detti che inducono un vero ripiegamento della proteina ed Esistono poi le sciap Peron che in aiutano semplicemente la proteina a ripiegarsi difetti degli chaperon portano a patologie da accumulo come la mucolipidosi Infatti il deficit di funzione delle proteine lisosomi dovuto a uno scorretto ripiegamento porta all'accumulo di
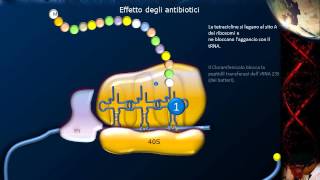
muco mucine e lipidi con comparsa precoce di lineamenti grossolani nel viso anomalie ossee Oltre a un ritardo psicomotorio tutto questo per un anormale ripiegamento della proteina Ovviamente la traduzione non avviene ad opera di un solo ribosoma ma un singolo RNA messaggero si attacca molto più di un ribosoma si forma quindi una conformazione chiamata polisoma in cui diversi ribosomi sono stati diversi del processo di traduzione un argomento strettamente legato alla traduzione è l'effetto dei farmaci antibiotici molti antibiotici Infatti agiscono bloccando la traduzione nei batteri le tetracicline per esempio bloccano il legame tra il sito a e
il tRNA il cloramfenicolo invece blocca la la peptidil trasferi dell RNA 23s dei batteri equivalente del 28s dei dei ribosomi eucarioti la streptomicina causa invece una errata lettura del codice ad opera della subunità minore infine l'eritromicina blocca il fattore di allungamento numero due va detto che questo stesso metodo di induzione della morte cellulare è utilizzato anche da alcuni batteri la tossina difterica per esempio ha lo stesso effetto dell'eritropoiesi sia secondo la via citoplasmatica la via escretoria prevede il RER il Golgi e dopo a seconda dei segnali di indirizzamento secondario si va all'esterno o verso i
lisosomi o verso la membrana tutto questo è mediato dalla srp Signal Recognition protein una proteina riconoscente ilale che sia sul reticolo endoplasmatico rugoso e riconosce il segnale di indirizzamento primario cioè una determinata sequenza all'interno dell RNA messaggero l'altra via è quella citoplasmatica cioè la trascrizione avviene adopera dei ribosomi liberi nel citoplasma il segnale di indirizzamento primario è la prima porzione della proteina appena sintetizzata dal RER la oltre la proteina matura è presente quindi questo segnale di indirizzamento formato da una regione positiva una regione idrofoba e una regione polare la molecola srp è fatta per riconoscere
questo segnale e legare il ribosoma che sta traducendo al RER Dopodiché l'intera proteina viene sintetizzata all'interno del Lume del RER grazie a una proteina canale se questo segnale non è presente le proteine vengono tradotte da RNA ribosomiali presenti nel citoplasma il destino delle proteine come abbiamo già visto nelle lezioni precedenti può seguire varie vie o una via secretoria o una via costitutiva o essere immessi all'interno dei perossisomi dei lisosomi o entrare nei mitocondri la distruzione delle proteine È un processo molto interessante e molto finemente regolato le proteine di base durano da alcuni minuti fino a
parecchi giorni vengono distrutte per vari motivi evitare l'accumulo delle proteine anomale che causa una serie di patologie chiamate malattie da accumulo evitare l'accumulo di quelle normali divenute inutili e infine favorire il riciclo degli aminoacidi la distruzione può seguire o la via lisosoma o la via ubiquitin dipendente La via lisos somale è una semplice distruzione Grazie all'auto fagocitosi la via ubiquitin dipendente Prevede invece che alla proteina vengano legate quattro molecole di ubiquitina queste vengono riconosciute da un complesso enzimatico chiamato proteasoma che ha la funzione di tagliare effettuare quindi molti tagli proteolitici per ridurre la proteina in
piccoli frammenti peptidici tutto ciò che abbiamo visto finora è possibile grazie al codice genetico e alle sue caratteristiche il codice genetico È un codice che viene letto a triplette cioè ogni parola dotata di senso deve avere tre basi non ha sovrapposizione Cioè non esiste un modo di leggerlo anomalo Non può essere spostata laa cornice di lettura ogni tripletta va Letta all'interno della tripletta stessa non è che una lettera viene Letta con la tripletta successiva il codice non ha punteggiatura è tutto attaccato Il codice è degenerato poiché esistono più triplette che possono Codificare per uno stesso
aminoacido questa caratteristica è chiamata anche ridondanza il codice è quasi universale sono solo due o tre le forme di vita che presentano un codice genetico lievemente diverso ma in tutte le forme di vita o quasi una stessa tripletta significa uno stesso amminoacido ovviamente il codice avrà dei segnali di inizio e fine dove il segnale di inizio è la tripletta a che codifica per la meteon i segnali finali sono corrisposti dalla tripletta uag uaa oppure Uga queste triplette dette non senso indicano il termine della trascrizione della traduzione il codice può presentare dei nucleotidi insoliti come Linosa
e presenta anche un fenomeno chiamato vacillamento dell'anticonformismo oltre all' inosina esistono anche altre basi come la uridina la metilguanosina la ribo idina eccetera sono basi molto particolari vengono incorporate a causa di piccole variazioni avvenute durante l'evoluzione sono rare Tuttavia l'ultima caratteristica del codice genetico È la ridondanza che però è dovuta al vacillamento dell'anticonformismo diverse triplette possono Codificare per uno stesso amminoacido questo avviene perché mentre la tripletta sull' RNA messaggero è lineare sull' RNA transfer la tripletta si trova su un'ansa quindi le tre basi divergono Perciò le prime due basi si legheranno bene l'ultima non si
legherà quasi quindi che All'ultimo posto ci sia una guanina un' adenina o qualsiasi altra base Il risultato non cambierà per cui triplette diverse potranno Codificare per uno stesso aminoacido perché riusciranno a legare l'rna transfer che presenta omologia tra le prime due basi ma l'ultima base diversa il legame avviene lo stesso perché a causa della forma curvilinea dell'anza dell'anticonformismo