[Music] hello and welcome back to immunology the war is over episode seven the compliment cascade when i first pictured this series in my mind i set aside only one episode for compliment and disease but when i actually sat down to create it i quickly realized that one episode was not going to be enough and so in this episode we're going to take a deep dive into the compliment cascade but then we'll follow on from there with some additional episodes which will focus on the complement regulators and some complement mediated conditions so stay tuned for those
okay so the compliment cascade in this episode i'm going to present compliment to you in two ways firstly we'll just recap on the basics and then we'll do a deep dive so depending on what you need for your studies the basics might be enough for you and you might want to stop there but i also wanted to present the deep dive version because complement is an area where the literature is exploding and the role of complement in various disease states is expanding across multiple specialties so as a doctor regardless of which field you're in understanding
complement is likely to come in handy in your future practice so take what you need from this episode if basic is enough stop there if you want a deep dive then keep going until the end and what's super fun about this episode is it's going to be handwritten i've provided you with a handy worksheet which you can download or you can just grab some blank paper and some colorful pens or highlighters and you can scribble along with me as we go so grab what you need and let's jump right in so let's start with the
basic overview which we touched on back in episode five complement is an incredible part of our innate immune system it's a proteolytic cascade which generates options to increase phagocytosis anaphyla toxins to attract immune cells to the area and the membrane attack complex which punches holes in target cells causing them to die and this goal of complement can be achieved via three different pathways the classical pathway the lectin pathway and the alternative pathway the classical pathway is predominantly triggered by immune complexes so an antibody stuck to the surface of a pathogen could spark this off but
equally the classical pathway can be triggered by damaged host tissue as well and the lectin pathway is pretty similar to the classical pathway but it's triggered slightly differently being sparked off by carbohydrate sequences on the surface of pathogens or by those same sequences on damaged cells and finally the alternative pathway which is the sassiest of the pathways this is always ticking over in the background and will activate quickly when it comes into contact with foreign cell surfaces and whilst these pathways are described separately they are not entirely exclusive they each have a tendency to feed
into and amplify each other but regardless of which pathway is sparked off first these three triggering pathways converge on the production of c3 converters then c5 converters and finally the formation of the membrane attack complex which punches holes in cells causing them to die and along the way the cascade produces anaphylatoxins and opsins anaphylotoxins c3a and c5a attract immune cells to the area and then promote the function of those immune cells when they arrive opsinins such as c3b and inactivated c3b promote phagocytosis of the cell they are attached to and there are some drugs which
have been designed to interrupt this cascade top of the list is echolusimab echolizumab is a monoclonal antibody which blocks c5 and so blocks the terminal pathway of complement this drug is one of the most expensive drugs in the world and is primarily used to treat two rare conditions for which there are no other effective treatments available namely atypical hus and peroxesmonocterno hemoglobin urea or pnh another complement blocking drug that's shown up in clinical trials recently is the oral c5 inhibitor avacapan this has recently been trialled as a steroid sparing agent in anchor vasculitis okay so
that's the basic overview done if your compliment cup is now full and you want to stop there that is absolutely fine but be sure to tune in again soon for the bonus episodes where compliment and disease come together i think you'll find them really useful for your exams now for those of you who want to stay for the compliment deep dive get ready to draw out the most beautiful compliment cascade you have ever seen as we draw this out we'll be building on the flowchart diagram that we just made but we're also going to place
these complement molecules on the cell surface so you can visualize what is actually happening and where it is happening so we can demystify this little cascade once and for all so let's start by unpacking the classical pathway this is triggered by immune complexes but it also interacts with dead cells and acute phase proteins to understand this pathway we must first be introduced to the c1 complex the c1 complex is composed of c1q r and s isn't it delightful when the alphabet is used to keep track of things so these molecules come together to make the
c1 complex and it almost looks like a little bunch of flowers the c1q molecules form the stems and the flower heads and the c1r and c1s molecules decorate the base of the bouquet but this is not a benign bunch of flowers it's a potent little pattern recognition molecule and i'm pretty sure it would feel moderately patronized at being likened to a bunch of flowers anyway these flower heads of the c1 complex are binding sites which recognize igg and igm immune complexes as well as exposed structures on damaged cells when c1 cube binds to something it
cleaves and activates c1r which in turn cleaves and activates c1s and c1s will now spark off the entire complement cascade by cleaving c2 and c4 now something you'll notice as we move through this tutorial is that as complement factors are cleaved they divide into two parts part a and part b again solid use of the alphabet here as a general rule part b wants to bind and belong to a team whilst part a would rather be alone you'll see why i'm telling you this in a second so c2 and c4 are cleaved into c2a c2b
c4a and c4b c4b and c2b want to belong to a team so they buddy up with each other on the cell surface and their a counterparts fly away i must say i couldn't find anything in the literature which assigned a particular function to c2a or c4a molecules so we'll just call them byproducts for now but c4b and c2b team up to make a bigger molecule c4b c2b which is as c3 converters now just a little side note historically older resources denote this as c4b c2a but the current modern terminology is c4b2b which let's face it
is much easier to work with because we have our little rule that part b wants to bind and belong to a team once the c3 converters establishes itself on the cell surface we are in business safety converters now cleaves c3 into c3a and c3b c3a is an anaphyllotoxin which will now go off and promote inflammation but c3b again wants to bind and belong so c3b will now join forces with the c3 converters molecule c4b2b to become c4b 2b 3b which is otherwise known as a c5 converters and c5 convertes as you would imagine cleave c5
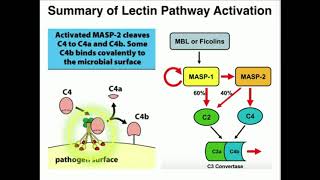
into you guessed it c5a and c5b c5a fulfills its role as an anaphyllotoxin and c5b again more of a team player joins up with c6 7 8 and 9 to form the membrane attack complex easy okay so that's the classical pathway done and dusted now let's shift our attention to the lectin pathway which is actually very similar to the classical pathway but with a few new immune characters in the mix now i don't know about you but the lectin pathway was something i misunderstood right from the start i knew there were molecules on the surface
of microbes which sparked everything off but i didn't understand the spark itself and the spark is fascinating the spark begins with something known as lectins lectins are pattern recognition molecules which bind to particular carbohydrate groups and when they do this they spark complement and the carbohydrate groups that are recognized by lectins are not only present on bacteria but also on viruses and fungi so lectins have a broad reach when it comes to identifying pathogens and whilst these carbohydrate groups are expressed on the surface of microbes they're not a feature of healthy human tissue but in
saying that if we have cell damage those same carbohydrate groups can be exposed and so just like the classical pathway the lectin pathway also has a role in clearing out damaged cells from the body and we have a few of these lectins whizzing around inside us the most famous of which is mbl mannose binding lectin mbl binds to a variety of bacteria and viruses including hiv but interestingly when it comes to fungal infections mbl binds to mannin which is a key component of the fungal cell wall it's part of what makes a fungus a fungus
and so it goes without saying that mbl has significant antifungal properties and whilst mbl is by far the most famous lectin we have we also have plenty of others such as the colectins and ficolins so lectins are pattern recognition molecules which can trigger the complement cascade but the true spark for complement is actually generated by what those lectins are attached to something known as masp mbl associated cerium protease as you can imagine this serine protease is in the business of cleaving proteins in other words masp is the true complement firestarter in the lectin pathway it
sparks everything off and when we draw this on our diagram what you'll notice is that this masp lectin complex looks very similar to the c1 qrs complex and indeed this is another potent little bunch of flowers we have the little flower heads otherwise known as carbohydrate recognition domains which when they bind signal downstream to mass which will in turn become activated and spark off the whole complement cascade and masp does this by cleaving c2 and c4 and so sparks off complement in the exact same way that we just saw in the classical pathway now as
with all things in the human body and particularly the immune system it's actually more complicated than that there are different types of mass and they interact with each other so mass 1 activates mass 2 which sparks off the lectin pathway we just described but mass 1 also activates mass 3 which is thought to promote the alternative pathway by activating factor d and of course we'll be discussing the alternative pathway shortly so that's the lectin pathway lectins are pattern recognition molecules connected to a potent spark called masp when they bind to particular carbohydrate motifs they will
recruit complement to annihilate organisms and or remove damaged tissue okay so now we have the classical and lectin pathways pretty much sorted now let's switch gears and get to know a real sassy part of complement the alternative pathway the alternative pathway has sas in spades it's a law unto itself and requires a significant amount of policing to keep it under control and in getting to know the alternative pathway we must first become acquainted with some brand new characters introducing factor b factor d and prepared in these are exclusive to the alternative pathway and for the
purpose of this tutorial we are going to use some wordplay to help us remember what these molecules actually do so b is for binding d is for dividing and p preparedn is for promotion and this will make a lot more sense in a tech when we see these characters in action but before we do that i just wanted to highlight another unique feature of the alternative pathway that gives it particular sas and sets it apart from the other pathways and this is the fact that the alternative pathway is always taking over in the background ready
for action so unlike the classical and lectin pathways which rely on specific pattern recognition molecules to get things going the alternative pathway does not require such a molecule and can be spontaneously activated of its own accords and in fact this is happening continuously at low levels all the time the so-called tick-over mechanism but this is confusing right what on earth do we mean when we say the alternative pathway is ticking over in the background let's unpack that now when we say the alternative pathway is ticking over what we really mean is that c3 is spontaneously
and continuously dividing into c3a and c3b and all it needs to do this is water so c3 and the presence of water happily divides into c3a and c3 watery b now these molecules are short-lived so they're always ticking over but there's not a lot of them around in any one moment but under the right circumstances these do have the potential to spark off the entire complement cascade so let's check out the alternative pathway in all of its cascading glory starting with the spontaneously generated c3 watery b this binds to factor b inner plasma and b
is for binding when factor b binds to activated c3 factor d comes along and divides it d is for divide factor d divides or cleaves factor b into b a and b b b a flies away whilst b b remains bound to c three b generating c3b bb which is a c3 converters capable of cleaving and activating c3 now this c3 converters is referred to as the fluid phase c3 converters because it's generated in the plasma but it will then try to establish itself on the surface of cells in this initial form it's actually pretty
unstable and prone to breakdown by complement regulators but this molecular complex can be strengthened by preparedn p is for promotion and prepared and binds to this molecule to become c3b bbp which is a much more stable version of c3 converters which resists breakdown by complement regulators and so remains active on the cell surface for longer so the plot thickens you can see now that we actually have two different versions of c3 converters depending on whether you start the cascade with the lectin classical or alternative pathways and similarly there are two different versions of c5 converters
in the alternative pathway another c3b is added to c3 converters to make c5 converters like so and so what becomes clear is the importance of c3b c3b is a part of the c3 converters in the alternative pathway but it's also a key part of c5 converters in both pathways and so c3b is a key component of the complement cascade at large but it also helps to establish a positive feedback loop whenever a c3 converters is generated it makes more c3b which in turn makes more c3 converters and so on and so forth but not only
do these molecules increase the production of each other they can also be added together to form c5 converters which will in turn lead to the formation of c5a and the membrane attack complex and so this cycle of c3b and c3 converters promoting each other and indeed the rest of the cascade is known as the amplification loop and whilst the amplification loop technically belongs to the alternative pathway it will be sparked by the classical and lectin pathways too so every time complement is activated you can expect to see this amplification loop in motion and this amplification
loop is potent sort of like adding gasoline to a burning flame you'd want to be careful not to add too much or things could quickly get out of hand and the same is true for complement we'll see in the next episode that there are a number of complement regulators many of whom make it their mission to interrupt this amplification loop by deactivating c3b and destabilizing c3 converters so ultimately the amplification loop is the balance between two competing cycles the formation of c3b and c3 converters as well as the breakdown of these molecules now we said
before that the alternative pathway doesn't have a particular trigger or pattern recognition molecule and it's always ticking over in the background spontaneously but that doesn't make perfect sense either sure it's always there in the background at low levels but how does it then ramp up to the full-blown cascade and this ultimately comes down to c3b binding to a surface it can thrive on so c3b is always being generated in the background and c3b is an opsonin which will bind to the surface of cells in its nearby vicinity so when c3b is produced it will absolutely
bind nearby cells including healthy cells but our body cells are fully aware of this issue and they use complement regulators on their surface to deactivate this molecule in a jiffy however fc3b happens to land on the surface of a bacterial cell which doesn't have any complement regulators it will be allowed to establish itself and thrive and this becomes the spark for the alternative pathway when c3b is allowed to thrive on the cell surface it will recruit all of its buddies from the alternative pathway so factor b and preparedn and in doing so generate c3 converters
and set up the amplification loop that we saw before which in turn will spark off a full-blown complement attack on that cell so that's the main way that we go from the alternative pathway ticking over in the background to sparking the complement cascade and you'd have to agree it's pretty clever but that's not the only way the alternative pathway can be activated and whilst technically pattern recognition molecules are not essential for the alternative pathway to happen there are a few molecules which can help things along for example preparedn when it binds to a microbial surface
can actually recruit complement to that location so as well as being able to stabilize c3 converters on the cell surface it may actually spark things off in the first place another molecule which has the potential to do this is p selecting which is secreted by endothelial cells and platelets and the magic of the alternative pathway doesn't stop there get this prepared in is not made in the liver it's actually made inside earth white blood cells monocytes macrophages and t cells can secrete preparedn into our bloodstream but other white cells such as neutrophils and mast cells
actually store preparedn in their granules and this is where it gets really good the complement cascade generates c5a an anaphylotoxin which attracts neutrophils to the area when the neutrophils arrive they release their granules which just happened to contain properdin which we now know can spark complement from the cell surface and can also stabilize c3 converters thereby promoting that amplification loop which will make more c5a more neutrophils will come to the area spew out prepared in and so on and so forth isn't that incredible and it's just one of the many examples of teamwork that takes
place inside our immune system and i won't go into detail on this but it's important to acknowledge that complement doesn't just intermingle with our innate immune cells it also influences our adaptive immune system as well or b cells and t cells for example b cells have a particular complement receptor on their surface cr2 which sits very close to the b cell receptor when a b cell comes across its matching antigen it will scan the nearby surface for complement using this receptor if a b cell finds complement together with its antigen it will lower the threshold
for activation of that b cell so when complement is there you'll need fewer antigens to get that b cell excited and present that antigen to t cells so that was the complement cascade in all its glory and now we have this fabulous diagram which summarizes the whole thing now whatever you do hold on to this diagram because you're gonna need it for the next episode when we see those complement regulators in action i'll see you in there bye [Music] you